
Как проверить микросхему на работоспособность мультиметром
При работе с электронными схемами часто требуется проверить исправность микросхем и ее составных частей, не выпаивая при этом их из платы. Для этой цели существуют несколько методов определения, начиная с визуального осмотра, заканчивая прозвоном с помощью специальных приборов. Наиболее надежной и доступной является проверка с использованием мультиметра.
Что такое мультиметр?
Мультиметр — это универсальное комбинированное измерительное устройство, которое объединяет функции нескольких измерительных устройств, то есть измеряет практически все показатели цепи. Самый маленький набор функций мультиметра — это измерение напряжения, тока и сопротивления. Однако современные производители не останавливаются на достигнутом, а вместо этого добавляют ряд функций, таких как емкостное измерение конденсаторов, частоты тока, проверка диодов (измерение падения напряжения на pn-переходе), звуковых датчиков, измерений температуры и измерения определенных параметров транзистора, встроенный генератор низких частот и многое другое.
Мультиметр может быть:
- Аналоговый. В данном типе приборов присутствует индикатор, который имеет несколько шкал (по одной на каждый вид измерения).
- Цифровой. Наиболее привычный вариант с цифровым табло. Показывает более точные значения. Имеет большее распространение по сравнению с аналоговым.
Устройство микросхемы
В составе микросхемы встречаются радиоэлементы, которые проверяются различными способами.
Конденсаторы, резисторы и диоды
Мультиметром можно проверить работоспособность конденсатора микросхемы, подключив прибор к его выходам. В очень короткий период времени значение сопротивления, отображаемое на устройстве, должно увеличиться с нескольких единиц до бесконечности. При изменении положения щупа также следует обратить внимание на это изменение.
Чтобы узнать, работает ли резистор в цепи должным образом, необходимо определить его сопротивление. Значение этого атрибута должно быть больше нуля, но не бесконечно большим. Если показатель на дисплее прибора не равен нулю или бесконечен во время теста, резистор работает нормально.
Если показатель на дисплее прибора не равен нулю или бесконечен во время теста, резистор работает нормально.
Процесс проверки диодов не очень сложен. Сначала необходимо определить сопротивление между катодом и анодом в одном порядке, затем изменить положение черного и красного проводов устройства. Работоспособность диода будет указываться стремлением к бесконечности числа, отображаемого на экране.
Индукционные катушки, тиристоры и стабилитроны
Чтобы проверить катушку на наличие неисправностей, также может понадобиться мультиметр. Если провод в мотке где-нибудь оборвется, устройство обязательно подаст сигнал. Все, что нужно сделать, чтобы проверить катушку, — это измерить ее сопротивление: оно не должно быть бесконечным. Стоит помнить, что не все доступные сегодня мультиметры могут проверять индуктивность.
Если необходимо определить, исправен ли такой компонент в микросхеме, как тиристор, необходимо выполнить следующие шаги:
- Сначала подключить красный провод к аноду, а черный — к катоду.
 Сразу после этого на экране устройства отображается информация, указывающая, что сопротивление стремится к бесконечности.
Сразу после этого на экране устройства отображается информация, указывающая, что сопротивление стремится к бесконечности. - Подсоединить контрольный электрод к аноду и наблюдать, как сопротивление уменьшается от бесконечности до нескольких единиц.
- После завершения процесса анод и электрод можно отсоединить друг от друга. В результате сопротивление, отображаемое на экране мультиметра, должно оставаться неизменным, равным нескольким Ом.
- Если во время теста все показатели в норме, то тиристор работает нормально и неисправностей нет.
 Сразу после этого на экране устройства отображается информация, указывающая, что сопротивление стремится к бесконечности.
Сразу после этого на экране устройства отображается информация, указывающая, что сопротивление стремится к бесконечности.Шлейф
Прозвонок шлейфа:
- Устанавливается режим измерения на мультиметре.
- Нужно проверить режим прозвона. Для проверки достаточно того, чтобы контакты щупа соприкоснулись. Если все в порядке, мультиметр подаст звуковой сигнал. В случае отсутствия звукового сопровождения нужно поменять прибор или заняться его ремонтом.
- Приклеить конец шлейфа к столу.
- Поместить красный щуп мультиметра на первый конец шлейфа и первый контакт.
- Поставить черный щуп на второй контакт и другой конец шлейфа.
Важно! Кабель состоит из тонких медных проводов, которые легко ломаются, поэтому шлейф не должен сгибаться.
Проверка микросхемы
Сложность проверки во многом зависит не только от метода, но и от устройства и особенностей конструкции микросхем. В конце концов, эти детали электронных вычислительных устройств, хотя и имеют одинаковые принципы построения, часто сильно отличаются друг от друга.
Например:
- Самый простой способ проверки — метод, относящийся к серии «КР142». Они имеют только три выхода, поэтому, когда какое-либо напряжение подается на один из входов, на выходе может использоваться контрольное устройство. После этого можно сразу сделать выводы о состоянии элемента.
- Более сложными типами являются «K155», «K176». Чтобы проверить их, необходимо использовать модуль с источником тока с определенным индикатором напряжения, который специально выбран для микросхемы. Характер проверки такой же, как и в первом варианте: просто подается напряжение на вход и проверяется выходной контакт с помощью мультиметра.
- Если необходимо выполнить более сложные тесты, которые не подходят для тестирования с помощью простого мультиметра, придется использовать специальный тестер цепи. Эти устройства могут быть изготовлены отдельно или приобретены в продаже. Тестеры могут помочь определить, работает ли конкретный узел цепи правильно. Как правило, данные, полученные во время теста, отображаются на экране устройства.
 Характер проверки такой же, как и в первом варианте: просто подается напряжение на вход и проверяется выходной контакт с помощью мультиметра.
Характер проверки такой же, как и в первом варианте: просто подается напряжение на вход и проверяется выходной контакт с помощью мультиметра.Важно! Напряжение, подаваемое на микросхему (микроконтроллер), не должно превышать нормальное значение или, наоборот, быть ниже требуемого уровня. Предварительная проверка может быть проведена на специально подготовленной испытательной доске.
Испытание микросхемыПроверка стабилизатора
Электронные компоненты, такие как стабилитроны, выглядят как диоды, но их использование в радиотехнике несколько иное.
Весь процесс похож на то, как проверяют диод. Это можно сделать в режиме тестирования резистора или диода с использованием обычного мультиметра. Как и диод, работающий стабилитрон может проводить ток в одном направлении.
Как проверить микросхему мультиметром
Первое и самое важное правило: можно проверять только полностью отключенную цепь, ни при каких обстоятельствах нельзя подключаться к проводам под напряжением.
Микросхема с помощью мультиметра проверяется по следующему алгоритму:
1. Устанавливается щуп в разъемы мультиметра:
— Красный штекер щупа в гнездо VΩmA
— Черный щуп в разъеме COM
2. Устройство включается поворотом регулятора, выбирается нужный режим, отмеченный нужным условным знаком. После этого на экране устройства должны отображаться цифры.
Устройство включается поворотом регулятора, выбирается нужный режим, отмеченный нужным условным знаком. После этого на экране устройства должны отображаться цифры.
3. Проверяется правильность работы мультиметра. Это делается путем соприкосновением контактов датчика . Если прибор работает нормально, то будет слышен звуковой сигнал, а на экране появится значение, близкое к нулю.
Как проверить работоспособность радиодеталей внешним осмотром
Внешний осмотр платы проводится в случаях, когда под рукой нет никаких приборов. Надежность этого способа не так велика. Если внимательно присмотреться к каждому элементу, есть вероятность обнаружить видимые дефекты. Например, это может быть сгоревший контакт или физическое повреждение Такой метод проверки устраняет необходимость в специальном оборудовании с мультиметром. Если дефекты видны невооруженным глазом, никакое оборудование не может быть использовано.
Важно! В противном случае все же придется прибегнуть к помощи специального оборудования.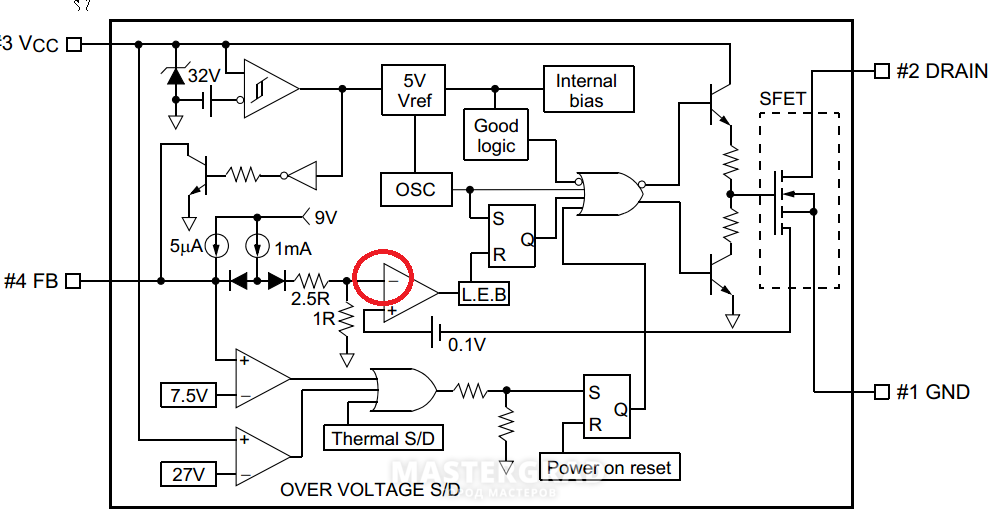
Меры безопасности
При использовании мультиметра необходимо строго соблюдать следующие правила электробезопасности:
- Нельзя применять мультиметр во влажной среде.
- Запрещается изменять режим работы и предел измерения в течение процесса.
- Измерение параметров, превышающих высший предел измерения прибора, запрещено.
- Запрещено включать в работу мультиметр с неисправным измерительным щупом.
Часто для проведения ремонтных и монтажных работ в радиоэлектронике требуется проверить работоспособность элементов платы. Выпаять и проверить каждый из них отдельно не представляется возможным, поэтому нужно знать, как проверить микросхему мультиметром, не выпаивая. Мультиметровая проверка будет наилучшим выбором. Это универсальный прибор, который прост в работе и доступен большинству пользователей.
Как проверить микросхему на работоспособность | Элементарно
Прежде чем проверять любую микросхему на работоспособность, необходимо знать и понимать ее устройство, хотя бы приблизительно. Это нужно для того, чтобы заранее представлять себе, какие сигналы или напряжения ожидать от исправной микросхемы на ее выводах.
Это нужно для того, чтобы заранее представлять себе, какие сигналы или напряжения ожидать от исправной микросхемы на ее выводах.
Лучше всего для проверки конкретной микросхемы собрать хотя бы на макетной плате схему для ее тестирования, — это в том случае, если микросхема новая или уже выпаяна.
Вообще, если устройство микросхемы известно, то в некоторых ситуациях ее можно проверить даже не выпаивая с платы, на которой она установлена, просто измерив сигналы на ножках при помощи мультиметра или осциллографа. Тогда наличие или отсутствие сигнала либо искаженная форма импульса сразу покажут, что — к чему.
Внешний осмотр микросхемы
Допустим что микросхема все еще установлена на плате и выпаивать ее сразу нежелательно. Прежде чем подавать питание на плату, внимательно осмотрите микросхему со всех сторон. Быть может есть очевидные физические признаки ее неисправности: трещина на корпусе, обгоревший или отпавший вывод, короткое замыкание между ножками из-за попадания куска провда (и такое бывет), горелые обвесные компоненты и т. д. Если при осмотре никаких поврежднеий не выявлено, можно идти дальше.
д. Если при осмотре никаких поврежднеий не выявлено, можно идти дальше.
Если к текущему моменту на плату подано питание, то можно аккуратно (с соблюдением техники безопасности!) приступать к дальнейшей проверке микросхемы.
Проверка выводов питания
Первым делом диагностируют цепи питания микросхемы. Это можно проделать при помощи вольтметра (мультиметра). Уточнить выводы питания известной микросхемы очень легко — достаточно заглянуть в документацию (datasheet) на нее. Плюс положительного питания обозначаетя в даташите как VCC+, отрицательное питание VCC-, общий провод имеет обозначение GND.
Итак, минусовой щуп мультиметра устанавливается на общий провод — упирается в минусовой вывод микросхемы, а плюсовой (красный) щуп мультиметра — на соответствующую ножку питания. Если напряжение соответствует норме для микросхемы, значит питание подается как надо, следовательно цепи питания всего устройства исправны.
Если же напряжение питания не в норме, значит необходимо далее проверить саму цепь питания, хотя бы предворительно отпаяв ее от микросхемы. Если цепи питания работают нормально без микросхемы, занчит проблема в микросхеме, и в худшем случае ее действительно придется менять. Если же проблема в цепях питания, значит скорее всего необходимо ремонтировать их (конденсатор, стабилизатор и т. д.).
Если цепи питания работают нормально без микросхемы, занчит проблема в микросхеме, и в худшем случае ее действительно придется менять. Если же проблема в цепях питания, значит скорее всего необходимо ремонтировать их (конденсатор, стабилизатор и т. д.).
Проверка источника опорного напряжения
Далее проверяют все известные выводы микросхемы. Например, можно начать с измерения напряжения на выводе встроенного в микросхему источника опорного напряжения Vref, нормальное значение которого указано в документации. На этом выводе должно быть постоянное напряжение определенного значения относительно общего провода. Если оно меньше или сильно больше, занчит внутри микросхемы или в обвесных компонентах что-то не так, и следует продолжить диагностику.
Проверка времязадающих цепей
Если на микросхеме есть какая-нибудь RC-цепь, то на ней, как правило, в рабочем режиме должны наблюдаться пилообразные колебания. На этом этапе опять же полезным будет обратиться к даташиту, чтобы понять где находится данная цепь если она предусмотрена, и на какой ножке должны быть колебания.
Проверка осуществляется осциллографом. Общий его щуп цепляется на минус питания, а измерительный — на соответствующий вывод микросхемы. Если колебания есть и их форма приемлема — все в порядке, можно идти дальше. Если колебаний нет, то скорее всего проблема в микросхеме или в обвесных времязадающих компонентах.
Проверка сигнальных выводов
Наконец, проверяют сигнальные выводы (выходы) микросхемы. Если микросхема управляет каким-то ключом или следующим блоком на схеме, то на соответствующих выходах (или хотя бы на одном выходе, если он единственный) микросхемы должны присутствовать правильные сигналы. Посмотрите в даташите, к каким выводам должны подходить управляемые цепи.
Проверьте осциллографом данные выводы тем же путем, как проверяли RC-цепь. Если сигнал нормальный и значительно не искажен по сравнению с нормальной формой, значит все в порядке. Если сигнал отсутствует или сильно искажен, скорее всего микросхема повреждена, и ее следует заменить, предварительно проверив управляемую цепь, ведь в действительности она может оказаться причиной выхода микросхемы из строя.
КАК ПРОВЕРИТЬ МИКРОСХЕМУ СТАБИЛИЗАТОР
Понадобилось собрать входные стабилизирующие цепи по питанию для устройства на основе микроконтроллера PIC16F628 стабильно работающего при напряжении от 5 вольт. Это не сложно. Взял интегральную микросхему PJ7805 и на её основе в соответствии со схемой из даташита сделал. Подал напряжение и на выходе получил 4,9 вольта. Всего скорей, что этого вполне достаточно, но упрямство, замешанное на педантичности, взяло верх.
Достал коробушку с интегральными стабилизаторами и вознамерился перемерить все соответствующего достоинства. А чтобы вдруг не ошибиться даже соответствующую схемку выложил перед собой. Однако энтузиазм закончился уже на первом же компоненте. Этот «ёжик без ручек, без ножек» из соединительных проводов с крокодилами желал жить своей жизнью и воли радиолюбителя подчинялся с большим трудом. Да к тому же проверяемый стабилизатор на выходе показал 4,86 вольта, чем поверг мой оптимизм в уныние.
Нет тут нужно что-то более существенное, например какой-то пусть и простой но, тем не менее, пробник что ли.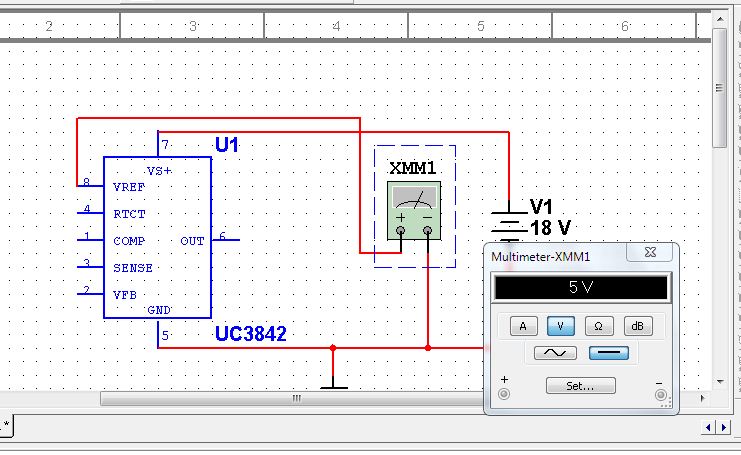 Забил в поисковик яндекса и получил то, что видите на фото «Комплекс контроля интегральных стабилизаторов напряжения». Ну, это не для средних радиолюбительских умов. Стало ясно, что велосипед придётся изобретать.
Забил в поисковик яндекса и получил то, что видите на фото «Комплекс контроля интегральных стабилизаторов напряжения». Ну, это не для средних радиолюбительских умов. Стало ясно, что велосипед придётся изобретать.
Схема испытателя КРЕН
Составленная схема явно уступает верхней картинке, ну тут уж ничего не поделаешь, что можем. Конденсатор С1 устраняет генерацию при скачкообразном включении входного напряжения, С2 служит для защиты от переходных помеховых импульсов. Их ёмкость решил взять 100 мкФ. Вольтаж в соответствии с напряжением проверяемого стабилизатора. Ставить конденсаторы как можно ближе к корпусу интегрального стабилизатора. Диод VD1 1N4148 не позволит конденсатору на выходе стабилизатора разрядится через него после выключения (это чревато выходом стабилизатора из строя). U Вх. интегрального стабилизатора должно быть выше U Вых. минимум на 2,5 вольта. Нагрузку подбирать так же в соответствии с возможностями тестируемого стабилизатора.
На роль корпуса был выбран самодельный вариант оборудованный контактными штырями для соединения с мультиметром (минус в гнездо «сom», плюс в «V»). В качестве соединительного элемента выводов проверяемого компонента со схемой можно приспособить вот такой тройной штыревой контакт. В мою задачу входит проверка трёхвыводных интегральных стабилизаторов рассчитанных на напряжение не более 12 вольт поэтому в схему поставлю два конденсатора 100 мкф х 16 В. Диод согласно схемы.
В качестве соединительного элемента выводов проверяемого компонента со схемой можно приспособить вот такой тройной штыревой контакт. В мою задачу входит проверка трёхвыводных интегральных стабилизаторов рассчитанных на напряжение не более 12 вольт поэтому в схему поставлю два конденсатора 100 мкф х 16 В. Диод согласно схемы.
В просверленные точно в соответствии с диаметром штыревых контактов отверстия их и вставляем, с внутренней стороны надеваем на каждый штырь по соответствующей (махонькой) металлической шайбочке, смочив активным флюсом и плотно прижав припаиваем каждую шайбу к соответствующему штырю не допуская соединения пар штырь – шайба между собой. Для этого шайбы нужно подточить, центральную с обеих сторон, крайние с одной. Отверстия по месту установки нужно
именно просверлить, если проколоть шилом образуется внутренняя неровность краёв отверстия и ровно + плотно установить шайбу не выйдет. Штыри, для прочности, также обязательно должны находится на общем твёрдом основании из диэлектрика.
Контактные площадки образованные местом пайки штырей и шайб становятся местом установки компонентов схемы. Получается компактно, также выполняется рекомендация минимального расстояния конденсаторов от выводов проверяемого интегрального стабилизатора. С соединительными проводами всё просто, главное взять их соответствующего цвета (для «+» красный, для «-» чёрный) и никакой путаницы не будет.
Подумав, установил кнопку включения нажимного действия, поставлена в разрыв плюсового (красного) провода на входе питания. Всё таки это удобство из разряда необходимых. Тройной штыревой контакт понадобилось «доработать» — немного согнуть, тут так, либо один раз подогнать контакты под выводы компонентов, либо перед каждым соединением ножки стабилизаторов гнуть под контакты.
Пробник – приставка к мультиметру готов. Вставляю в соответствующие гнёзда мультиметра штыри пробника, предел измерения выставляю 20 вольт постоянного напряжения, провода подвода электрического тока подсоединяю к лабораторному блоку питания в соответствии с их расплюсовкой, устанавливаю для проверки стабилизатор (попался на 10 вольт), выставляю соответственно на БП напряжение 15 вольт и нажимаю кнопку включения на пробнике. Устройство сработало, на дисплее 9,91 В. Далее в течении минуты разобрался со всеми трёхвыводными стабилизаторами на напряжение до 12 вольт включительно. Несколько, из числа бережно хранимых, оказались негодными.
Устройство сработало, на дисплее 9,91 В. Далее в течении минуты разобрался со всеми трёхвыводными стабилизаторами на напряжение до 12 вольт включительно. Несколько, из числа бережно хранимых, оказались негодными.
Итого
Давно понятно, что вот такие простенькие пробники – приставки в радиолюбительском деле так же необходимы, как и весьма серьёзные измерительные приборы, но вот делать их (возиться с их изготовлением) попросту лень, а напрасно, и понимание этого приходит каждый раз когда это простенькое устройство всё же было собрано и оказало неоценимую помощь в творческих начинаниях. Автор — Babay iz Barnaula.
Форум
Форум по обсуждению материала КАК ПРОВЕРИТЬ МИКРОСХЕМУ СТАБИЛИЗАТОР
Проверка работоспособности шим-контроллера.
Шим-контроллер считают «сердцем» источников питания, но предварительно нужно проверить и другие компоненты блока питания выполнив стандартную последовательность действий по ремонту блока питания (БП):
1) В выключенном состоянии источник внимательно осмотреть (особое внимание обратить на состояние всех электролитических конденсаторов — они не должны быть вздуты).
2) Проверить исправность предохранителя и элементов входного фильтра БП.
3) Прозвонить на короткое замыкание или обрыв диоды выпрямительного моста (эту операцию, как и многие другие, можно выполнить, не выпаивая диоды из платы). При этом в остальных случаях надо быть уверенным, что проверяемая цепь не шунтируется обмотками трансформатора или резистором (в подозрительных случаях, элемент схемы необходимо выпаивать и проверять отдельно).
4) Проверить исправность выходных цепей: электролитических конденсаторов низкочастотных фильтров, выпрямительных диодов и диодных сборок.
5) Проверить силовые транзисторы высокочастотного преобразователя и транзисторов каскада управления. Обязательно проверить возвратные диоды, включенные параллельно электродам коллектор-эмиттер силовых транзисторов.
Эти действия, дают положительный результат в обнаружении только следствия неработоспособности всего блока, но причина неисправности в большинстве случаев находится гораздо глубже. Например, неисправность силовых транзисторов может быть следствием: неисправности цепей схемы защиты и контроля, нарушения цепи обратной связи, неисправности ШИМ-преобразователя, выхода из строя демпфирующих RC-цепочек или, межвитковый пробой в силовом трансформаторе. Поэтому, если удается найти неисправный элемент, то желательно пройти все этапы проверок, перечисленные выше (т. к. предохранитель сам по себе никогда не сгорает, а пробитый диод в выходном выпрямителе становится причиной «смерти» ещё и силовых транзисторов высокочастотного преобразователя).
Например, неисправность силовых транзисторов может быть следствием: неисправности цепей схемы защиты и контроля, нарушения цепи обратной связи, неисправности ШИМ-преобразователя, выхода из строя демпфирующих RC-цепочек или, межвитковый пробой в силовом трансформаторе. Поэтому, если удается найти неисправный элемент, то желательно пройти все этапы проверок, перечисленные выше (т. к. предохранитель сам по себе никогда не сгорает, а пробитый диод в выходном выпрямителе становится причиной «смерти» ещё и силовых транзисторов высокочастотного преобразователя).
В качестве шим-контроллера («сердца» источников питания) долгое время использовали микросхему TL494, а затем и ее аналоги (MB3759, KA7500B … KA3511, SG6105 и др.). Проверку работоспособности такой микросхемы, например, TL494 (рис. 1) можно произвести, не включая блок питания. При этом микросхему необходимо запитать от внешнего источника напряжением +9В..+20В. Напряжение подается на вывод 12 относительно выв. 7 — желательно через маломощный выпрямительный диод.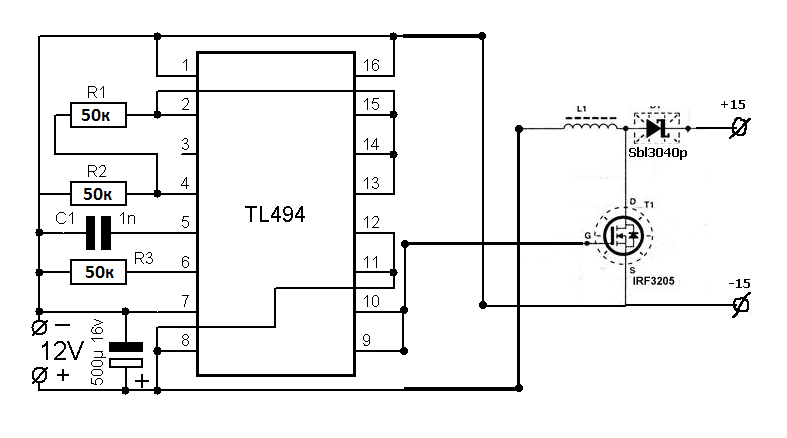 Все измерения тоже должны проводиться относительно выв. 7. При подаче питания на микросхему контролируем напряжение на выв. 5. Оно должно быть +5В (±5%) и быть стабильным при изменении напряжения питания на выв. 12 В пределах +9В..+20В. В противном случае не исправен внутренний стабилизатор напряжения микросхемы. Далее осциллографом смотрим напряжение на выв. 5. Оно должно быть пилообразной формы амплитудой 3,2 В (рис. 2). Если сигнал отсутствует или иной формы, то проверить целостность конденсатора и резистора, подключенных к выв. 5 и выв. 6, соответственно. В случае исправности этих элементов микросхему необходимо заменить. После этого проверяем наличие управляющих сигналов на выходе микросхемы (выв. 8 и выв. 11). Они должны соответствовать осциллограммам, приведенным на рис. 2. Отсутствие этих сигналов так же говорит о неисправности микросхемы. В случае успешного прохождения испытаний микросхема считается исправной.
Все измерения тоже должны проводиться относительно выв. 7. При подаче питания на микросхему контролируем напряжение на выв. 5. Оно должно быть +5В (±5%) и быть стабильным при изменении напряжения питания на выв. 12 В пределах +9В..+20В. В противном случае не исправен внутренний стабилизатор напряжения микросхемы. Далее осциллографом смотрим напряжение на выв. 5. Оно должно быть пилообразной формы амплитудой 3,2 В (рис. 2). Если сигнал отсутствует или иной формы, то проверить целостность конденсатора и резистора, подключенных к выв. 5 и выв. 6, соответственно. В случае исправности этих элементов микросхему необходимо заменить. После этого проверяем наличие управляющих сигналов на выходе микросхемы (выв. 8 и выв. 11). Они должны соответствовать осциллограммам, приведенным на рис. 2. Отсутствие этих сигналов так же говорит о неисправности микросхемы. В случае успешного прохождения испытаний микросхема считается исправной.
Рис. 1
Рис. 2
Как проверить микросхему на стойкость ко внешним воздействиям
Анализ защиты от внешнего воздействия
В условиях быстро развивающейся отрасли ИТ-технологий перед производителями и пользователями защищенных информационных систем встают вопросы защиты и противодействия несанкционированному доступу, требующие всё новых решений.
Сегодня хакеры используют всё более изощрённые методы получения несанкционированного доступа. Определение уязвимости, связанной с ошибками в проектировании аппаратных средств, с ошибками в программных кодах, с ошибками в результате случайно или намеренно вносимых сбоев в работу аппаратных средств, ЭМ-воздействие на аппаратные средства и т.п. — все это связано с оценкой стойкости электронных устройств ко внешним воздействиям. От правильности выбранного программно-аппаратного решения зависит защищенность информации от внешних атак хакеров.
Компания Riscure (Нидерланды) — общепризнанный лидер в отрасли производства оборудования для проверки стойкости аппаратных средств и программного кода ко внешним воздействиям. Наряду с этим компания Riscure располагает собственной лабораторией оказывающей услуги по тестированию защищенности микросхем и встроенных систем на соответствие отраслевым стандартам или техническому заданию заказчика.
Оборудование для анализа устойчивости ко внешним воздействиям
Для анализа устойчивости электронных компонентов и систем, таких как микропроцессоры, смарт-карты, сим-карты, встроенные системы (system-on-chip) и т. п., используется два вида оборудования:
п., используется два вида оборудования:
- Оборудование для анализа устойчивости к атакам по сторонним или побочным каналам SCA
- Оборудование для анализа внесением искажений FI.
Riscure Inspector — оборудование для оценки и анализа уязвимости чипов ко внешним воздействиям
Линейка оборудования Riscure Inspector
— это мощная и гибкая исследовательская платформа, которая включает в себя интегрированный набор аппаратного и программного обеспечения для оценки и анализа уязвимостей чипов и систем к пассивным видам атак по побочному каналу SCA (SIDE CHANNEL ANALYSIS) и активным видам атак по ошибкам вычислений методом внесения искажений FI (FAULT INJECTION).
Система Inspector SCA
Система Inspector SCA предназначена для оценки устойчивости устройств к атакам по побочному каналу (SIDE CHANNEL ATTACKS). В ней применяются методы анализа энергопотребления устройств – DPA (Differential Power Analysis), CPA (Correlation Power Analysis), а также набор тестовых процедур TVLA (Test Vector Leakage Assessment).
- DPA — дифференциальный анализ энергопотребления. Основан на статистическом анализе результатов множества измерений данных энергопотребления устройства, обусловленных физической реализацией механизма алгоритма, устранения влияния шумов и погрешностей измерения.
- CPA – корреляционный анализ энергопотребления. Основан на вычислении вектора коррелирующих с ключом значений энергопотребления с пиком во время его выполнения.
- TVLA – оценка утечек при помощи тестовых векторов. Данная методика включает набор тестов, позволяющих определить потенциальные уязвимости систем по нескольким параметрам – энергопотреблению, радио- и электромагнитному излучениям и т.д.
Данные методы позволяют аналитикам оценивать возможность преобразования физических данных в бинарные, следствием чего может быть извлечение ключа и оказание несанкционированного воздействия на систему злоумышленниками.
Система Inspector FI
Оборудование Riscure Inspector FI
Система Inspector FI предназначена для оценки устойчивости к атакам методом внесения искажений (FAULT INJECTION) и включает в себя модуль дифференциального анализа искажений DFA (Differential Fault Analysis) для оценки устойчивости к атакам на ошибки вычислений для алгоритмов шифрования DES , AES
и RSA. Предлагает все функции для тестирования технологий применяемых в смарт-картах. С помощью Inspector FI пользователь может проверить, можно ли извлечь ключ, вызвав ошибки в операциях чипа, обойти проверку, такую как аутентификация или состояние жизненного цикла, или изменить программный поток кода на чипе.
Предлагает все функции для тестирования технологий применяемых в смарт-картах. С помощью Inspector FI пользователь может проверить, можно ли извлечь ключ, вызвав ошибки в операциях чипа, обойти проверку, такую как аутентификация или состояние жизненного цикла, или изменить программный поток кода на чипе.
Используемые аппаратные компоненты обеспечивают точные, надежные и, следовательно, предсказуемые, вносимые сбои тактовой частоты и напряжения, а также усовершенствованные оптические атаки с помощью специально разработанного лазерного оборудования. Атаки, связанные с внесением искажений, также известные как атаки с возмущением, изменяют нормальное поведение микросхемы, вызывая эксплуатационную ошибку.
С помощью Inspector FI пользователь может проверить, можно ли извлечь ключ, вызвав ошибки в операциях чипа, обойти проверку, такую как аутентификация или состояние жизненного цикла, или изменить программный поток кода на чипе. Инспектор FI — Fault Injection — предлагает все функции для проведения тестирования внесением искажений для технологии смарт-карт.
Единая платформа Riscure Inspector
Обе модификации систем Inspector SCA и Inspector FI могут быть объединены в единой платформе Inspector SCA/FI. Оборудование Inspector SCA и FI
позволяет разработчикам и производителям устройств исследовать их защищенность и подбирать адекватные механизмы внешней и внутренней защиты от хакерских атак.
Если Вас заинтересовало оборудование для проверки на стойкость ко внешним воздействиям, обращайтесь в нашу компанию. Наши специалисты расскажут подробно о методах проверки и сориентируют Вас по ценам на продукцию. Присылайте свои вопросы на электронную почту: [email protected]
Проверка радиодеталей мультиметром
Серия статей известного автора множества радиолюбительских публикаций Дригалкина В.В. для начинающих радиолюбителей
Доброго дня уважаемые радиолюбители!
Приветствую вас на сайте “Радиолюбитель“

Без измерительного прибора Вам не обойтись, т.к. придется проверять сопротивление резисторов, напряжения и тока в разных цепях конструкций. Измерительный прибор, в народе – омметр, авометр (ампер-вольт-омметр) , тестер или мультиметр (от английского multimeter – измерительный прибор, объединяющий в себе несколько функций) – должен иметь каждый. Сейчас большой популярностью пользуются цифровые приборы. Они многофункциональные и сравнительно не дорогие . Ранее в качестве измерительного прибора широко пользовались аналоговыми тестерами со стрелочным индикатором (см. Рис. 1).
Не все начинающие знают, что омметром можно проверять почти все радиоэлементы : резисторы, конденсаторы, катушки индуктивности, трансформаторы, диоды, тиристоры, транзисторы, некоторые микросхемы. В авометре омметр образован внутренним источником тока (сухим элементом или батареей), стрелочным прибором и набором резисторов, которые переключаются при изменении пределов измерения. Сопротивления резисторов подобраны таким образом, чтобы при коротком замыкании клемм омметра стрелка прибора отклонилась вправо до последнего деления шкалы. Это деление соответствует нулевому значению измеряемого сопротивления. Когда же клеммы омметра разомкнуты, стрелка прибора стоит напротив левого крайнего деления шкалы, которое обозначено значком бесконечно большого сопротивления. Если к клеммам омметра подключено какое-то сопротивление, стрелка показывает промежуточное значение между нулем и бесконечностью, и отсчет производится по оцифровке шкалы. В связи с тем, что шкалы омметров выполняются в логарифмическом масштабе, края шкалы получаются сжатыми. Поэтому наибольшая точность измерения соответствует положению стрелки в средней, растянутой части шкалы. Таким образом, если стрелка прибора оказывается у края шкалы, в сжатой ее части, для повышения точности отсчета следует переключить омметр на другой предел измерения.
Сопротивления резисторов подобраны таким образом, чтобы при коротком замыкании клемм омметра стрелка прибора отклонилась вправо до последнего деления шкалы. Это деление соответствует нулевому значению измеряемого сопротивления. Когда же клеммы омметра разомкнуты, стрелка прибора стоит напротив левого крайнего деления шкалы, которое обозначено значком бесконечно большого сопротивления. Если к клеммам омметра подключено какое-то сопротивление, стрелка показывает промежуточное значение между нулем и бесконечностью, и отсчет производится по оцифровке шкалы. В связи с тем, что шкалы омметров выполняются в логарифмическом масштабе, края шкалы получаются сжатыми. Поэтому наибольшая точность измерения соответствует положению стрелки в средней, растянутой части шкалы. Таким образом, если стрелка прибора оказывается у края шкалы, в сжатой ее части, для повышения точности отсчета следует переключить омметр на другой предел измерения.
Омметр производит измерение сопротивления, подключенного к его клеммам, путем измерения постоянного тока, протекающего в измерительной цепи. Поэтому к измеряемому сопротивлению прикладывается постоянное напряжение от встроенного в омметр источника. В связи с тем, что некоторые детали обладают разными сопротивлениями постоянному току в зависимости от полярности приложенного напряжения , для грамотного использования омметра необходимо знать, какая из клемм омметра соединена с плюсом источника тока, а какая – с минусом. В паспорте авометра эти сведения обычно не указаны, и их нужно определить самостоятельно . Это можно сделать либо по схеме авометра, либо экспериментально с помощью какого-либо дополнительного вольтметра или исправного диода любого типа. Щупы омметра подключают к вольтметру так, чтобы стрелка вольтметра отклонялась вправо от нуля. Тогда тот щуп, который подключен к плюсу вольтметра, будет также плюсовым, а второй – минусовым. При использовании в этих целях диода два раза измеряют его сопротивление; сначала произвольно подключая к диоду щупы, а второй раз – наоборот. За основу берется то измерение, при котором показания омметра получаются меньшими. При этом щуп, подключенный к аноду диода, будет плюсовым, а щуп, подключенный к катоду диода, – минусовым.
При проверке исправности того или иного радиоэлемента возможны две различные ситуации: либо проверке подлежит изолированный, отдельный элемент, либо элемент, впаянный в какое-то устройство. Нужно учесть, что, за редкими исключениями, проверка элемента, впаянного в схему, не получится полноценной, при такой проверке возможны грубые ошибки. Они связаны с тем, что параллельно контролируемому элементу в схеме могут оказаться подключены другие элементы, и омметр будет измерять не сопротивление проверяемого элемента, а сопротивление параллельного соединения его с другими элементами. Оценить возможность достоверной оценки исправности контролируемого элемента схемы можно путем изучения этой схемы, проверяя, какие другие элементы к нему подключены и как они могут повлиять на результат измерения. Если такую оценку произвести затруднительно или невозможно, следует отпаять от остальной схемы хотя бы один из двух выводов контролируемого элемента и только после этого производить его проверку. При этом также не следует забывать и о том, что тело человека также обладает некоторым сопротивлением, зависящим от влажности кожной поверхности и от других факторов. Поэтому при пользовании омметром во избежание появления ошибки измерения нельзя касаться пальцами обоих выводов проверяемого элемента.
Проверка резисторов
Проверка постоянных резисторов производится омметром путем измерения их сопротивления и сравнения с номинальным значением, которое указано на самом резисторе и на принципиальной схеме аппарата. При измерении сопротивления резистора полярность подключения к нему омметра не имеет значения. Необходимо помнить, что действительное сопротивление резистора может отличаться по сравнению с номинальным на величину допуска. Поэтому, например, если проверяется резистор с номинальным сопротивлением 100 кОм и допуском ±10%, действительное сопротивление такого резистора может лежать в пределах от 90 до 110 кОм. Кроме того, сам омметр обладает определенной погрешностью измерения (обычно порядка 10%) . Таким образом, при отклонении фактически измеренного сопротивления на 20% от номинального значения резистор следует считать исправным.
1. Вообще то, где какой щуп указано на корпусе любого авометра.
2. Если он не оборван, то исправен и всегда может пригодится.
При проверке переменных резисторов измеряется сопротивление между крайними выводами, которое должно соответствовать номинальному значению с учетом допуска и погрешности измерения, а также необходимо измерять сопротивление между каждым из крайних выводов и средним выводом. Эти сопротивления при вращении оси из одного крайнего положения в другое должны плавно, без скачков изменяться от нуля до номинального значения. При проверке переменного резистора, впаянного в схему, два из его трех выводов необходимо выпаивать. Если переменный резистор имеет дополнительные отводы, допустимо, чтобы только один вывод оставался припаянным к остальной части схемы.
Проверка конденсаторов
В принципе конденсаторы могут иметь следующие дефекты: обрыв, пробой и повышенная утечка. Пробой конденсатора характеризуется наличием между его выводами короткого замыкания, то есть нулевого сопротивления. Поэтому пробитый конденсатор любого типа легко обнаруживается омметром путем проверки сопротивления между его выводами. Конденсатор не пропускает постоянного тока, его сопротивление постоянному току, которое измеряется омметром, должно быть бесконечно велико. Однако это оказывается справедливо лишь для идеального конденсатора. В действительности между обкладками конденсатора всегда имеется какой-то диэлектрик, обладающий конечным значением сопротивления, которое называется сопротивлением утечки. Его-то и измеряют омметром. В зависимости от используемого в конденсаторе диэлектрика устанавливаются критерии исправности по величине сопротивления утечки. Слюдяные, керамические, пленочные, бумажные, стеклянные и воздушные конденсаторы имеют очень большое сопротивление утечки, и при их проверке омметр должен показывать бесконечно большое сопротивление . Однако имеется большая группа конденсаторов, сопротивление утечки которых сравнительно невелико. К ней относятся все полярные конденсаторы, которые рассчитаны на определенную полярность приложенного к ним напряжения, и эта полярность указывается на их корпусах. При измерении сопротивления утечки этой группы конденсаторов необходимо соблюдать полярность подключения омметра (плюсовой вывод омметра должен присоединяться к плюсовому выводу конденсатора), в противном случае результат измерения будет неверным. К этой группе конденсаторов в первую очередь относятся все электролитические конденсаторы и оксидно-полупроводниковые. Сопротивление утечки исправных конденсаторов этой группы должно быть не менее 100 кОм, остальных не менее 1 МОм. При проверке конденсаторов большой емкости нужно учесть, что при подключении омметра к конденсатору, если он не был заряжен, начинается его зарядка, и стрелка омметра делает бросок в сторону нулевого значения шкалы. По мере зарядки стрелка движется в сторону увеличения сопротивлений. Чем больше емкость конденсатора, тем медленнее движется стрелка. Отсчет сопротивления утечки следует производить только после того, как она практически остановится. При проверке конденсаторов емкостью порядка 1000 мкФ на это может потребоваться несколько минут. Внутренний обрыв или частичная потеря емкости конденсатором не могут быть обнаружены омметром, для этого необходим прибор, позволяющий измерять емкость конденсатора. Однако обрыв конденсатора емкостью более 0,2 мкФ может быть обнаружен омметром по отсутствию начального скачка стрелки во время зарядки . Следует заметить, что повторная проверка конденсатора на обрыв по отсутствию начального скачка стрелки может производиться только после снятия заряда, для чего выводы конденсатора нужно замкнуть на короткое время.
Конденсаторы переменной емкости проверяются омметром на отсутствие замыканий. Для этого омметр подключается к каждой секции агрегата и медленно поворачивается ось из одного крайнего положения в другое. Омметр должен показывать бесконечно большое сопротивление в любом положении оси.
Проверка катушек индуктивности
При проверке катушек индуктивности омметром контролируется только отсутствие в них обрыва. Сопротивление однослойных катушек должно быть равно нулю, сопротивление многослойных катушек близко к нулю. Иногда в паспортных данных аппарата указывается сопротивление многослойных катушек постоянному току и на его величину можно ориентироваться при их проверке. При обрыве катушки омметр показывает бесконечно большое сопротивление. Если катушка имеет отвод, нужно проверить обе секции катушки, подключая омметр сначала к одному из крайних выводов катушки и к ее отводу, а затем – ко второму крайнему выводу и отводу.
Проверка низкочастотных дросселей и трансформаторов
Как правило, в паспортных данных аппаратуры или в инструкциях по ее ремонту указываются значения сопротивлений обмоток постоянному току, которые можно использовать при проверке трансформаторов и дросселей. Обрыв обмотки фиксируется по бесконечно большому сопротивлению между ее выводами. Если же сопротивление значительно меньше номинального, это может указывать на наличие короткозамкнутых витков. Однако чаще всего короткозамкнутые витки возникают в небольшом количестве, когда происходит замыкание между соседними витками, и сопротивление обмотки изменяется незначительно. Для проверки отсутствия короткозамкнутых витков можно поступить следующим образом. У трансформатора выбирается обмотка с наибольшим количеством витков, к одному из выводов которой подключается омметр с помощью зажима “крокодил”. Ко второму выводу этой обмотки прикасаются слегка влажным пальцем левой руки. Держа металлический наконечник второго щупа омметра правой рукой, подключают его ко второму выводу обмотки, не отрывая от него пальца левой руки. Стрелка омметра отклоняется от своего начального положения, показывая сопротивление обмотки. Когда стрелка остановится, отводят правую руку с щупом от второго вывода обмотки. В момент разрыва цепи при исправном трансформаторе чувствуется легкий удар электрическим током, возникающей при разрыве цепи. В связи с тем, что энергия разряда мизерна, никакой опасности такая проверка не представляет. Омметр при этом нужно использовать на самом меньшем пределе измерения, который соответствует наибольшему току измерения.
Проверка диодов
Полупроводниковые диоды характеризуются резко нелинейной вольтамперной характеристикой. Поэтому их прямой и обратный токи при одинаковом приложенном напряжений различны. На этом основана проверка диодов омметром. Прямое сопротивление измеряется при подключении плюсового вывода омметра к аноду, а минусового вывода – к катоду диода. У пробитого диода прямое и обратное сопротивления равны нулю. Если диод оборван, оба сопротивления бесконечно велики.
Указать заранее значения прямого и обратного сопротивлений или их соотношение нельзя, так как они зависят от приложенного напряжения, а это напряжение у разных авометров и на разных пределах измерения различно. Тем не менее, у исправного диода обратное сопротивление должно быть больше прямого. Отношение обратного сопротивления к прямому у диодов, рассчитанных на низкие обратные напряжения, велико (может быть более 100). У диодов, рассчитанных на большие обратные напряжения, это отношение оказывается незначительным, так как обратное напряжение, приложенное к диоду омметром, мало по сравнению с тем обратным напряжением, на которое диод рассчитан. Методика проверки стабилитронов и варикапов не отличается от изложенной. Как известно, если к диоду приложено напряжение, равное нулю, ток диода также будет равен нулю. Для получения прямого тока необходимо приложить к диоду какое-то пороговое небольшое напряжение . Любой омметр обеспечивает приложение такого напряжения. Однако если соединено последовательно и согласно (в одну сторону) несколько диодов, пороговое напряжение, необходимое для отпирания всех диодов, увеличивается и может оказаться больше, чем напряжение на клеммах омметра. По этой причине измерить прямые напряжения диодных столбов или селеновых столбиков при помощи омметра оказывается невозможно.
Проверка тиристоров.
Неуправляемые тиристоры (динисторы) могут быть проверены таким же образом, как диоды, если напряжение отпирания динистора меньше напряжения на клеммах омметра. Если же оно больше, динистор при подключении омметра не отпирается и омметр в обоих направлениях показывает очень большое сопротивление. Тем не менее, если динистор пробит, омметр это регистрирует нулевыми показаниями прямого и обратного сопротивлений. Для проверки управляемых тиристоров (тринисторов) плюсовой вывод омметра подключается к аноду тринистора, а минусовой вывод – к катоду. Омметр при этом должен показывать очень большое сопротивление, почти равное бесконечному. Затем замыкают выводы анода и управляющего электрода тринистора, что должно приводить к резкому уменьшению сопротивления, так как тринистор отпирается. Если после этого отключить управляющий электрод от анода, не разрывая цепи, соединяющей анод тринистора с омметром, для многих типов тринисторов омметр будет продолжать показывать низкое сопротивление открытого тринистора. Это происходит в тех случаях, когда анодный ток тринистора оказывается больше так называемого тока удержания. Тринистор остается открытым обязательно, если анодный ток больше гарантированного тока удержания. Это требование является достаточным, но не необходимым. Отдельные экземпляры тринисторов одного и того же типа могут иметь значения тока удержания значительно меньше гарантированного. В этом случае тринистор при отключении управляющего электрода от анода остается открытым. Но если при этом тринистор запирается и омметр показывает большое сопротивление, нельзя считать , что тринистор неисправен.
Проверка транзисторов.
Эквивалентная схема биполярного транзистора представляет собой два диода, включенных навстречу один другому. Для p-n-р транзисторов эти эквивалентные диоды соединены катодами, а для n-p-п транзисторов – анодами. Таким образом, проверка транзистора омметром сводится к проверке обоих р-n переходов транзистора: коллектор-база и эмиттер-база. Для проверки прямого сопротивления переходов p-n-р транзистора минусовой вывод омметра подключается к базе, а плюсовой вывод омметра – поочередно к коллектору и эмиттеру. Для проверки обратного сопротивления переходов к базе подключается плюсовой вывод омметра. При проверке n-p-п транзисторов подключение производится наоборот: прямое сопротивление измеряется при соединении с базой плюсового вывода омметра, а обратное сопротивление – при соединении с базой минусового вывода. При пробое перехода его прямое и обратное сопротивления оказываются равными нулю. При обрыве перехода его прямое сопротивление бесконечно велико. У исправных маломощных транзисторов обратные сопротивления переходов во много раз больше их прямых сопротивлений. У мощных транзисторов это отношение не столь велико, тем не менее, омметр позволяет их различить. Из эквивалентной схемы биполярного транзистора вытекает, что с помощью омметра можно определить тип проводимости транзистора и назначение его выводов (цоколевку). Сначала определяют тип проводимости и находят вывод базы транзистора. Для этого один вывод омметра подключают к одному выводу транзистора, а другим выводом омметра
касаются поочередно двух других выводов транзистора. Затем первый вывод омметра подключают к другому выводу транзистора, а другим выводом омметра касаются свободных выводов транзистора. Затем первый вывод омметра подключают к третьему выводу транзистора, а другим выводом касаются остальных. После этого меняют местами выводы омметра и повторяют указанные измерения. Нужно найти такое подключение омметра, при котором подключение второго вывода омметра к каждому из двух выводов транзистора, не подключенных к первому выводу омметра, соответствует небольшому сопротивлению (оба перехода открыты). Тогда вывод транзистора, к которому подключен первый вывод омметра, является выводом базы. Если первый вывод омметра является плюсовым, значит, транзистор относится к n-p-п проводимости, если – минусовым, значит, – p-n-р проводимости. Теперь нужно определить, какой из двух оставшихся выводов транзистора является выводом коллектора. Для этого омметр подключается к этим двум выводам, база соединяется с плюсовым выводом омметра при n-p-п транзисторе или с минусовым выводом омметра при p-n-р транзисторе и замечается сопротивление, которое измеряется омметром. Затем выводы омметра меняются местами (база остается подключенной к тому же выводу омметра, что и ранее) и вновь замечается сопротивление по омметру. В том случае, когда сопротивление оказывается меньше, база была соединена с коллектором транзистора.
Проверка деталей цифровым мультиметром.
Главным отличием цифрового прибора от аналогового является то, что результаты измерения отображаются на жидкокристаллическом дисплее. К тому же цифровые мультиметры обладают более высокой точностью и отличаются простотой использования, т.к. не приходится разбираться во всех тонкостях градирования измерительной шкалы, как со стрелочными измерительными приборами.
Цифровой тестер (см. Рис. 1), как и аналоговый, имеет два щупа – черный и красный, и от двух до четырех гнезд. Черный вывод является общим (масса). Гнездо для общего вывода помечается как СОМ или просто “-” (минус), а сам вывод на конце часто имеет так называемый пкрокодильчикп, для того, чтобы при измерении можно было зацепить его за массу электронной схемы. Красный вывод вставляется в гнездо, помеченное символами напряжения – “V” или “+” (плюс).
Если Ваш прибор содержит более двух гнезд, например, как на Рис. 1, красный щуп вставляется в гнездо “VQmA”. Эта надпись говорит о том, что Вы можете измерять напряжение, сопротивление и небольшой ток – в миллиамперах. Гнездо, расположение немного выше, с маркировкой 10ADC говорит о том, что Вы можете измерять большой постоянный ток, но не выше 10А.
Переключатель мультиметра позволяет выбрать один из нескольких пределов для измерений.
Чтобы измерить постоянное напряжение выбираем режим DCV1, если переменное ACV, подключаем щупы и смотрим результат. При этом на шкале переключателя вы должны выбрать большее напряжение, чем измеряемое. Например, Вам необходимо измерить напряжение в электрической розетки. В вашем приборе шкала ACV состоит из двух параметров: 200 и 750 (это вольты). Значит, нужно установить стрелочку переключателя на параметр 750 и можно смело измерять напряжение.
1 DC – постоянный ток (Direct Current), AC – переменный ток (Alternating Current).
Ток измеряется последовательным включением мультиметра в электрическую цепь. Для примера можно взять обычную лампочку от карманного фонаря и подключить ее последовательно с прибором к адаптеру 5В. Корда по цепи пойдет ток и лампочка загорится, прибор покажет значение тока.
Сопротивление на приборе обозначается значком, немного похожим на наушники. Для измерения сопротивления резистор должен быть выпаян из электрической цепи хотя бы одним концом, чтобы быть уверенным в том, что никакие другие компоненты схемы не повлияют на результат. Подключаем щупы к двум концам резистора и сравниваем показания омметра со значением, которое указано на самом резисторе . Стоит учитывать и величину допуска (возможных отклонений от нормы), т.е. если по маркировке резистор на 200кОм и допуском ± 15%, его действительное сопротивление может быть в пределах 170-230кОм.
Проверяя переменные резисторы, измеряем сначала сопротивление между крайними выводами (должно соответствовать номиналу резистора), а затем подключив щуп мультиметра к среднему выводу, поочередно с каждым из крайних. При вращении оси переменного резистора, сопротивление должно изменяться плавно, от нуля до его максимального значения, в этом случае удобней использовать аналоговый мультиметр наблюдая за движением стрелки, чем за быстро меняющимися цифрами на жидкокристаллическом экране.
Для проверки диодов типовые приборы содержат специальный режим. В более дешевых тестерах можно воспользоваться режимом прозвонки. Тут все просто: в одну сторону диод звониться, а в другую – нет. Проверить диод можно и в режиме сопротивления. Для этого устанавливаем переключатель на 1к0м. При подключении красного вывода мультиметра к аноду диода, а черного к катоду, Вы увидите его прямое сопротивление, при обратном подключении сопротивление будет настолько высоко, что на данном пределе измерения вы не увидите ничего. Если диод пробит, его сопротивление в любую сторону будет равно нулю, если оборван, то в любую сторону сопротивление будет бесконечно большим.
Обычный биполярный транзистор представляет собой два диода, включенных навстречу один другому. Зная, как проверяются диоды, несложно проверить и такой транзистор. Стоит не забывать, что транзисторы бывают разных типов: у р-п-р условные диоды соединены катодами, у п-р-п – анодами. Для измерения прямого сопротивления транзисторных p-n-р переходов, минус мультиметра подключается к базе, а плюс поочередно к коллектору и эмиттеру. При измерении обратного опротивления меняем полярность. Для проверки транзисторов п-р-п типа делаем все наоборот. Если еще короче, то переходы база-коллектор и база-эмиттер в одну сторону должны прозваниваться, в другую – нет.
Для измерения у транзистора коэффициента усиления по току используем режим hEF, если он есть на Вашем приборе. Разъем, в который вставляют контакты транзистора для измерения hEF, не очень качественный практически во всех моделях тестеров и довольно глубоко посажен. То есть ножки транзистора до них иногда не достают. Как выход – вставьте одножильные провода и выводами транзистора касайтесь именно их.
На цифровых мультиметрах пределов измерений обычно больше, к тому же часто добавлены дополнительные функции, например, частотомер, измеритель емкости конденсаторов и даже датчик температуры. Но такими возможностями обладают более дорогие модели тестеров. Кроме того, в дорогих моделях отсутствует необходимость переключать шкалу измерения. Просто устанавливаете переключать на измерение емкости, сопротивления и т.д., и прибор показывает результат.
Для того, чтобы мультиметр не вышел из строя при измерениях напряжения или тока, особенно если их значение неизвестно, переключатель желательно установить на максимально возможный предел измерений, и только если показание при этом слишком мало, для получения более точного результата, переключайте мультиметр на предел ниже текущего.
Как проверить операционный усилитель. » Хабстаб
На днях купил в магазине операционный усилитель(ОУ) за 1.5$, пришёл домой, запаял, тишина. То что виноват в работоспособности схемы ОУ сомнений не было, поэтому выпаял купленный ОУ и решил проверить. Соединил инвертирующий вход с выходом, подал питание и напряжение на прямой вход(1V), исправный ОУ на выходе должен был выдать то, что подал ему на вход, собственно в этом и заключается проверка ОУ, а у меня на выходе ноль.Интересно, подумал тогда, либо перегрел его когда паял, что вряд ли, либо купил неисправный. Снова пошёл в магазин, купил ещё один, но решил проверить его перед тем как запаивать и о чудо, этот то же неисправный, но теперь его хоть можно вернуть продавцу, судя по всему, у него таких целая партия…
Но разбираться времени не было, пошёл в другой магазин и купил такой же ОУ, но уже за 4$, при покупке договорились, что если он не заработает то, принесу его обратно. Пришёл домой, проверил — работает, запаял — работает. Вывод из этого можно сделать следующий, после покупки детали, перед тем как её запаивать желательно проверить, а продавец, скорее всего, заказал партию этих ОУ с Китая и когда получил, не проверил, это и понятно когда у тебя целый магазин с радиодеталями проверять все устанешь.
К чему всё это писал, после этого поискал эти ОУ на али и когда нашёл их был приятно удивлён, на те деньги, которые потратил у себя в городе чтобы купить исправный ОУ(4$) в Китае можно было купить 5 штук, но они были в корпусе soic8, а имея негативный опыт, описанный выше, конечно же, хотелось их проверить когда они придут. Решить этот вопрос можно было несколькими способами, вытравить макетку, в которую можно было впаивать ОУ каждый раз, с другой стороны, чтобы не впаивать можно было просто прижимать ОУ к плате прищепкой, уже лучше, но есть вариант ещё интереснее, так как часто приходиться иметь дело с soic8, решил поискать ZIF адаптер soic8 – dip8, тогда можно будет собрать схему на breadboard, что значительно ускорит процесс.
В общем нашел такой переходник на али за 1.7$ и это с учётом доставки. Когда ОУ пришли, переходник был уже на руках, а так как у меня в арсенале есть генератор сигналов, то проверял их по схеме из даташита.
Что интересно, все заказанные ОУ оказались исправными.
soic8 — dip8 ZIF adapter
Определение микросхемы Merriam-Webster
mi · cro · cir · cuit | \ ˈMī-krō-ˌsər-kət \Определение микросхемы
Микросхема, активируемая ацетилхолином, управляет временной динамикой корковой активности
Kitt, C.A., Höhmann, C., Coyle, J.T. & Прайс, Д. Холинергическая иннервация структур переднего мозга мышей. J. Comp. Neurol. 341 , 117 (1994).
CAS PubMed Google Scholar
Метерат, Р., Кокс, С. и Эш, Дж. Клеточные основы неокортикальной активации: модуляция нервных колебаний базальным ядром и эндогенным ацетилхолином. J. Neurosci. 12 , 4701–4711 (1992).
CAS PubMed Google Scholar
Гоард М. и Дэн Ю. Активация базального переднего мозга усиливает кортикальное кодирование естественных сцен. Nat. Neurosci. 12 , 1444 (2009).
CAS PubMed PubMed Central Google Scholar
Харрис, К.Д. & Тиле, А. Состояние коры и внимание. Nat. Rev. Neurosci. 12 , 509–523 (2011).
CAS PubMed PubMed Central Google Scholar
Herrero, J.L. et al. Ацетилхолин через мускариновые рецепторы участвует в модуляции внимания в V1. Природа 454 , 1110 (2008).
CAS PubMed PubMed Central Google Scholar
Cohen, M.R. & Maunsell, J.H.R. Внимание улучшает производительность в первую очередь за счет уменьшения межнейронных корреляций. Nat. Neurosci. 12 , 1594–1600 (2009).
CAS PubMed PubMed Central Google Scholar
Тиле, А. Оптимизация обработки мозга. Nat. Neurosci. 12 , 1359–1360 (2009).
CAS PubMed Google Scholar
Li, C.Y., Poo, M.M. И Дэн Ю. Всплеск одиночного коркового нейрона изменяет глобальное состояние мозга. Наука 324 , 643–646 (2009).
CAS PubMed PubMed Central Google Scholar
Gentet, L.J., Avermann, M., Matyas, F., Staiger, J.F. & Petersen, C.C.H. Динамика мембранного потенциала ГАМКергических нейронов в бочкообразной коре головного мозга мышей. Нейрон 65 , 422–435 (2010).
CAS PubMed Google Scholar
Ренарт, А.и другие. Асинхронное состояние в корковых цепях. Наука 327 , 587–590 (2010).
CAS PubMed PubMed Central Google Scholar
King, P.D., Zylberberg, J. & DeWeese, M.R. Тормозящие интернейроны декоррелируют возбуждающие клетки, чтобы управлять формированием разреженного кода в модели пиков V1. J. Neurosci. 33 , 5475–5485 (2013).
CAS PubMed PubMed Central Google Scholar
Сиппи Т.& Юсте, Р. Декорреляционное действие торможения в неокортикальных сетях. J. Neurosci. 33 , 9813–9830 (2013).
CAS PubMed PubMed Central Google Scholar
Klausberger, T. et al. Активизация интернейронов гиппокампа, специфичная для состояния мозга и типа клеток in vivo . Природа 421 , 844–848 (2003).
CAS PubMed Google Scholar
Кардин, Дж.A. et al. Управление быстропротекающими клетками вызывает гамма-ритм и контролирует сенсорные реакции. Природа 459 , 663–667 (2009).
CAS PubMed PubMed Central Google Scholar
Arroyo, S., Bennett, C., Aziz, D., Brown, SP & Hestrin, S. Длительное дисинаптическое торможение в коре головного мозга, опосредованное медленным, не-α7 никотиновым возбуждением определенного подмножества корковых интернейронов . J. Neurosci. 32 , 3859–3864 (2012).
CAS PubMed PubMed Central Google Scholar
Xiang, Z., Huguenard, J.R. & Prince, D.A. Холинергическое переключение в тормозных сетях неокортекса. Наука 281 , 985–988 (1998).
CAS PubMed Google Scholar
Кавагути Ю. Селективная холинергическая модуляция подтипов ГАМКергических клеток коры. J. Neurophysiol. 78 , 1743–1747 (1997).
CAS PubMed Google Scholar
Fanselow, E.E., Richardson, K.A. И Коннорс, Б. Селективная, зависимая от состояния активация ингибирующих интернейронов, экспрессирующих соматостатин, в неокортексе мышей. J. Neurophysiol. 100 , 2640–2652 (2008).
PubMed PubMed Central Google Scholar
Сюй, Х., Jeong, H.-Y., Tremblay, R. & Rudy, B. Неокортикальные ГАМКергические интернейроны, экспрессирующие соматостатин, подавляют таламореципиентный слой 4. Neuron 77 , 155–167 (2013).
CAS PubMed PubMed Central Google Scholar
Алито, Х.Дж. и Дэн, Й. Модуляция активности неокортекса, специфичная для клеточного типа, посредством входного сигнала базального переднего мозга. Фронт. Syst. Neurosci. 6 , 79 (2012).
PubMed Google Scholar
Fu, Y. et al. Корковый контур для управления усилением поведенческим состоянием. Ячейка 156 , 1139–1152 (2014).
CAS PubMed PubMed Central Google Scholar
Arroyo, S., Bennett, C. & Hestrin, S. Никотиновая модуляция корковых цепей. Фронт. Нейронные схемы 8 , 30 (2014).
PubMed PubMed Central Google Scholar
Letzkus, J.J. и другие. Растормаживающая микросхема для обучения ассоциативному страху в слуховой коре. Природа 480 , 331–335 (2011).
CAS PubMed PubMed Central Google Scholar
Kalmbach, A. & Waters, J. Модуляция высокочастотных и низкочастотных компонентов кортикального локального поля потенциала через никотиновые и мускариновые рецепторы ацетилхолина у анестезированных мышей. J. Neurophysiol. 111 , 258–272 (2014).
CAS PubMed Google Scholar
Chen, N. et al. Специфическая для стимула пластичность в зрительной коре, обусловленная базальным ядром, опосредуется астроцитами. Proc. Natl. Акад. Sci. США 109 , 2832–2841 (2012).
Google Scholar
Пфеффер, К.К., Сюэ, М., Хе, М., Хуанг, З.Дж. & Сканциани, М. Ингибирование торможения в зрительной коре: логика связей между молекулярно различными интернейронами. Nat. Neurosci. 16 , 1068–1076 (2013).
CAS PubMed PubMed Central Google Scholar
Цзян, X., Wang, G., Lee, A.J., Stornetta, R.L. & Zhu, J.J. Организация двух новых корковых межнейронных цепей. Nat. Neurosci. 16 , 210–218 (2013).
CAS PubMed PubMed Central Google Scholar
Pi, H.-J. и другие. Корковые интернейроны, специализирующиеся на растормаживающем контроле. Природа 503 , 521–524 (2013).
CAS PubMed PubMed Central Google Scholar
Кругликов И. и Руди Б. Перисоматическое высвобождение ГАМК и таламокортикальная интеграция в возбуждающие клетки неокортекса регулируются нейромодуляторами. Нейрон 58 , 911–924 (2008).
CAS PubMed PubMed Central Google Scholar
Фино, Э. и Юсте, Р. Плотная ингибирующая связь в неокортексе. Нейрон 69 , 1188–1203 (2011).
CAS PubMed PubMed Central Google Scholar
Арамакис, В.Б., Бандровски, А.Э. и Эш, Дж.H. Мускариновое снижение ГАМКергических синаптических потенциалов приводит к растормаживанию АМРА / каинат-опосредованного ВПСП в слуховой коре. Brain Res. 758 , 107–117 (1997).
CAS PubMed Google Scholar
McCormick, D.A. И Принс, Д.А. Механизмы действия ацетилхолина в коре головного мозга морских свинок in vitro . J. Physiol. (Лондон) 375 , 169–194 (1986).
CAS Google Scholar
Giessel, A.J. И Сабатини, Б. Мускариновые рецепторы M1 усиливают синаптические потенциалы и приток кальция в дендритные шипы, ингибируя постсинаптические sk-каналы. Нейрон 68 , 936 (2010).
CAS PubMed PubMed Central Google Scholar
Бьюкенен, К.А., Петрович, М.М., Чемберлен, С.E.L., Marrion, N.V. & Mellor, J.R. Содействие долгосрочному усилению мускариновыми рецепторами m1 опосредуется ингибированием sk-каналов. Нейрон 68 , 948 (2010).
CAS PubMed PubMed Central Google Scholar
де Ла Роша, Дж., Дуарон, Б., Ши-Браун, Э., Йосич, К. и Рейес, А. Корреляция между последовательностями нервных импульсов увеличивается с увеличением скорости стрельбы. Природа 448 , 802–806 (2007).
CAS PubMed Google Scholar
Rudy, B., Fishell, G., Lee, S. & Hjerling-Leffler, J. Три группы интернейронов составляют почти 100% неокортикальных ГАМКергических нейронов. Dev. Neurobiol. 71 , 45–61 (2011).
PubMed PubMed Central Google Scholar
Адесник, Х., Брунс, В., Танигучи, Х., Хуанг, З.J. & Scanziani, M. Нейронная цепь для пространственного суммирования в зрительной коре. Природа 490 , 226–231 (2012).
CAS PubMed PubMed Central Google Scholar
Gentet, L.J. et al. Уникальные функциональные свойства ГАМКергических нейронов, экспрессирующих соматостатин, в коре головного мозга мышей. Nat. Neurosci. 15 , 607–612 (2012).
CAS PubMed Google Scholar
Марросу, Ф.и другие. Измерение микродиализом высвобождения ацетилхолина кортикальным слоем и гиппокампа во время цикла сна-бодрствования у свободно движущихся кошек. Brain Res. 671 , 329–332 (1995).
CAS PubMed Google Scholar
Poulet, J.F.A., Fernandez, L.M.J., Crochet, S. & Petersen, C.C.H. Таламический контроль корковых состояний. Nat. Neurosci. 15 , 370–372 (2012).
CAS PubMed Google Scholar
Харрис, К.D. Контроль состояния коры головного мозга сверху вниз. Нейрон 79 , 408–410 (2013).
CAS PubMed PubMed Central Google Scholar
Vanderwolf, C.H. И Стюарт, Д. Таламический контроль неокортикальной активации: критическая переоценка. Brain Res. Бык. 20 , 529–538 (1988).
CAS PubMed Google Scholar
Компте, А., Санчес-Вивес, М.В., Маккормик, Д.А. И Ван, X.-J. Клеточные и сетевые механизмы медленной колебательной активности (<1 Гц) и распространения волн в модели корковой сети. J. Neurophysiol. 89 , 2707–2725 (2003).
PubMed Google Scholar
Саката, С. и Харрис, К.Д. Ламинарная структура спонтанной и сенсорно-вызванной активности населения в слуховой коре. Нейрон 64 , 404–418 (2009).
CAS PubMed PubMed Central Google Scholar
McCormick, D.A. Действие нейротрансмиттеров в таламусе и коре головного мозга и их роль в нейромодуляции таламокортикальной активности. Прог. Neurobiol. 39 , 337 (1992).
CAS PubMed Google Scholar
Фрайз, П., Рейнольдс, Дж. Х., Рори, А. Э. и Дезимон, Р.Модуляция колебательной нейрональной синхронизации избирательным зрительным вниманием. Наука 291 , 1560–1563 (2001).
CAS Google Scholar
Робертс, М.Дж. и др. Ацетилхолин динамически контролирует пространственную интеграцию в первичной зрительной коре мартышек. J. Neurophysiol. 93 , 2062–2072 (2005).
CAS PubMed Google Scholar
Винье, W.Э. и Галлант, Дж. Л. Редкое кодирование и декорреляция в первичной зрительной коре при естественном зрении. Наука 287 , 1273–1276 (2000).
CAS PubMed Google Scholar
Гизельманн, М.А. и Тиле, А. Сравнение характеристик пространственной интеграции и подавления объемного звука при пиковой активности и потенциале локального поля у макак V1. евро. J. Neurosci. 28 , 447–459 (2008).
CAS PubMed Google Scholar
Müller, C.M. & Singer, W. Индуцированное ацетилхолином угнетение зрительной коры головного мозга кошки опосредуется ГАМКергическим механизмом. Brain Res. 487 , 335–342 (1989).
PubMed Google Scholar
Zhao, S. et al. Трансгенные мыши родопсина-2 со специфическим типом клеток для оптогенетического анализа функции нервных цепей. Nat. Методы 8 , 745–752 (2011).
CAS PubMed PubMed Central Google Scholar
Madisen, L. et al. Надежная и высокопроизводительная система Cre отчетов и характеристик для всего мозга мыши. Nat. Neurosci. 13 , 133–140 (2010).
CAS PubMed PubMed Central Google Scholar
Чоу, Б.Y. et al. Высокоэффективное генетически нацеленное оптическое нейронное подавление с помощью световых протонных насосов. Природа 463 , 98–102 (2010).
CAS PubMed PubMed Central Google Scholar
Wilson, N.R., Runyan, C.A., Wang, F.L. & Sur, M. Деление и вычитание различными корковыми тормозными сетями in vivo . Природа 488 , 343–348 (2012).
CAS PubMed PubMed Central Google Scholar
Рунян, К.A. et al. Характеристики ответа интернейронов, экспрессирующих парвальбумин, предполагают точную роль подтипов торможения в зрительной коре. Нейрон 67 , 847–857 (2010).
CAS PubMed PubMed Central Google Scholar
Kuhlman, S.J. И Хуанг, З.Дж. Мечение с высоким разрешением и функциональные манипуляции с конкретными типами нейронов в мозге мыши с помощью Cre-активированной экспрессии вирусного гена. PLoS One 3 , e2005 (2008 г.).
PubMed PubMed Central Google Scholar
Tamamaki, N. et al. Экспрессия зеленого флуоресцентного белка и совместная локализация с кальретинином, парвальбумином и соматостатином у мышей с нокаутом GAD67-GFP. J. Comp. Neurol. 467 , 60–79 (2003).
CAS PubMed Google Scholar
Pologruto, T.A., Sabatini, B.L. И Свобода, К.ScanImage: гибкое программное обеспечение для работы с лазерными сканирующими микроскопами. Biomed. Англ. Онлайн 2 , 13 (2003).
PubMed PubMed Central Google Scholar
Джоши, С. и Хокен, М.Дж. Запись оккстеклеточной области с произвольным участком in vivo : метод функциональной характеристики и маркировки нейронов у макака V1. J. Neurosci. Методы 156 , 37–49 (2006).
CAS PubMed Google Scholar
Кардин, Дж.A. et al. Направленная оптогенетическая стимуляция и регистрация нейронов in vivo с использованием специфической для клеточного типа экспрессии Channelrhodopsin-2. Nat. Protoc. 5 , 247–254 (2010).
CAS PubMed PubMed Central Google Scholar
Brainard, D.H. The Psychophysics Toolbox. Spat. Vis. 10 , 433 (1997).
CAS PubMed PubMed Central Google Scholar
Кальмбах, А., Хедрик, Т. и Уотерс, Дж. Селективная оптогенетическая стимуляция холинергических аксонов в неокортексе. J. Neurophysiol. 107 , 2008–2019 (2012).
CAS PubMed PubMed Central Google Scholar
фон Энгельгардт, Дж., Элиава, М., Мейер, А.Х., Розов, А. и Моньер, Х. Функциональная характеристика внутренних холинергических интернейронов в коре головного мозга. J. Neurosci. 27 , 5633–5642 (2007).
CAS PubMed PubMed Central Google Scholar
Tricoire, L. & Cea-Del Rio, C.A. Освещение холинергических микросхем неокортекса. J. Neurosci. 27 , 12119–12120 (2007).
CAS PubMed PubMed Central Google Scholar
ДеФелипе Дж. И Фариньяс И. Пирамидный нейрон коры головного мозга: морфологические и химические характеристики синаптических входов. Прог. Neurobiol. 39 , 563–607 (1992).
CAS PubMed Google Scholar
Connors, B.W. & Gutnick, M.J. Внутренние паттерны возбуждения различных нейронов неокортекса. Trends Neurosci. 13 , 99–104 (1990).
CAS PubMed Google Scholar
Prestori, F. et al. Стробирование долгосрочной потенциации никотиновыми ацетилхолиновыми рецепторами на стадии входа в мозжечок. PLoS ONE 8 , e64828 (2013).
CAS PubMed PubMed Central Google Scholar
Руководство по технологии гибридных микросхем — 2-е издание
Уесть собственная консалтинговая фирма AvanTeco, специализирующаяся на материалах и процессах для электроники. Он имеет степень бакалавра химии в Фордхэмском университете и докторскую степень по химии в Принстонском университете, где он был старшим научным сотрудником DuPont.Его области знаний включают материалы и процессы для электронных приложений, в первую очередь для систем высокой надежности, гибридных микросхем, печатных схем и других технологий межкомпонентной упаковки. Он является экспертом в области полимерных материалов, включая клеи, покрытия, герметики, изоляцию, надежность, основанную на режимах и механизмах отказов. Доктор Ликари имеет сорокалетнюю карьеру, посвященную изучению и развитию микроэлектронных материалов и процессов.
Среди заметных достижений на протяжении всей этой карьеры — проведение первых исследований надежности и использования клеев для прикрепления матриц для микросхем, которые он проводил в середине 1970-х — начале 1980-х годов, что позволило промышленности и правительству осознать разрушительное влияние следов количество ионных примесей в эпоксидных смолах.Он провел ранние исследовательские разработки по использованию паст для толстопленочных проводников из неблагородных металлов (Cu) в схемах из толстопленочной керамики. Он провел первые исследования по использованию парилена в качестве диэлектрического и пассивирующего покрытия для МОП-устройств и в качестве иммобилайзера частиц для гибридных микросхем. Он разработал первые фотоопределяемые толстопленочные пасты для проводников и резисторов, которые были предшественниками процесса Фоделя DuPont, на который он получил патент, выданный в Англии. И он разработал первое фотоотверждаемое эпоксидное покрытие с использованием катионного фотоинициирования, используя соль диазония в качестве каталитического агента (U.С. 3205157). Эта работа упоминается как новаторская в обзорной статье Дж. В. Кривелло «Открытие и разработка катионных фотоинициаторов на основе ониевых солей», J. Polymer Chemistry (1999)
Варианты подключения, которые позволяют и ограничивают нейронные вычисления в сенсорной микросхеме
Ниже мы прилагаем полные обзоры всех трех рецензентов. Мы надеемся, что вы сможете ответить на каждую критику в своей редакции. Мы хотели бы прояснить один пункт, касающийся «размера синапса» (первая критика рецензента 3).После дальнейшего обсуждения рецензентами рецензент 3 сделал следующие дополнительные комментарии: «Взаимосвязь между размером синапса, а также количеством синапсов и силой соединения является частью общей предпосылки исследований дрозофилы. большинство синапсов имеют одинаковый размер, так что сила пути действительно приближается к числу синапсов без ссылки на их размер, но даже в этом случае размер синапса редко сообщается и сравнивается между областями нейропиля.Чтобы продемонстрировать модульность размера синапсов, необходимо было бы измерить только ~ 40 синаптических профилей в нормальном поперечном сечении и 20 в продольном сечении, чтобы определить вариацию размеров контактов синапсов и сравнить с другими опубликованными отчетами. Это не только затронет важный компонент силы проводящих путей, но и признает важную проблему, которая может быть неочевидной для читателей, работающих с различными нервными системами ».
Мы согласны с тем, что размер синапсов является важным вопросом.Здесь есть два вопроса, на которые мы можем ответить с помощью нашего набора данных. Во-первых, насколько различается размер синапсов? Во-вторых, как вариации размера синапсов соотносятся с другими анатомическими вариациями?
Чтобы ответить на эти вопросы, мы случайным образом выбрали 10 ORN (5 слева и 5 от правой антенны) и вручную сегментировали все их синапсы на все 5 наших реконструированных PN (n T-образных стержня = 683, n профили пн = 1106). В каждом синапсе были измерены два признака: пресинаптический объем Т-образного стержня ORN и площадь постсинаптического контакта PN (PSCA) для всех присутствующих постсинаптических профилей PN.Чтобы повысить точность этих измерений, каждый объем Т-образного стержня и PSCA были независимо измерены тремя разными аннотаторами, а затем измерения разных аннотаторов были объединены после взвешивания измерений каждого аннотатора в соответствии с их общей погрешностью измерения по всему набору данных (относительная к другим аннотаторам).
Результаты нашего анализа размера синапсов теперь показаны в новом дополнении (Рисунок 1 — Приложение 6). В целом, мы обнаружили, что два показателя размера синапса (объем Т-образного стержня и площадь постсинаптического контакта) были значительно (хотя и слабо) коррелированы.Они также значительно коррелировали с количеством постсинаптических профилей на Т-образный стержень. Это означает, что должны быть механизмы, которые производят скоординированные вариации всех трех переменных на основе синапса за синапсом (объем Т-образного стержня, площадь постсинаптического контакта и количество постсинаптических профилей).
Тем не менее, когда мы сравнивали различные соединения ORN → PN (путем усреднения размера всех синапсов, принадлежащих одному и тому же соединению), мы обнаружили, что размер синапса был лишь незначительным источником вариации между соединениями (рисунок 1 — приложение к рисунку 6). .Основным источником различий между соединениями по-прежнему было количество синапсов на одно соединение (рис. 5). Другими словами, то, что отличает одно соединение от другого, — это прежде всего количество синапсов, которые оно содержит, а не размер этих синапсов.
Рецензент № 1:
Это отличная статья, объединяющая новые данные ЭМ-реконструкции с моделированием и предыдущими электрофизиологическими исследованиями, чтобы дать общее представление о взаимосвязи между структурой, физиологическими свойствами и функциями микросхем.Авт. Сосредотачиваются на реконструкции нейронов обонятельных рецепторов (ORNs) и проекционных нейронов (PNs), которые иннервируют одну из пятидесяти пар клубочков в антеннальной доле Drosophila. Помимо получения ценных количественных ультраструктурных данных о синаптической связности, авторы создали компартментальную модель, которая имитирует, как входы ORN интегрируются в PN, и использовали ее для анализа трех интересных аспектов своих данных: как PN компенсируют разницу в их количестве. (особенно интересно), почему ипси-латеральные ORNs обнаруживают более сильные связи с PN, чем контрлатеральные ORN, и каково функциональное значение вариаций в количестве синапсов между отдельными ORN → PN парами.Авторы также явно обсудили ограничения своего исследования (например, их модель не принимает во внимание тормозящий вход или вариации силы отдельных синапсов). Среди недавнего всплеска исследований ЭМ-реконструкции этот считается одним из лучших для интеграции структуры и физиологических свойств хорошо изученной модельной схемы, и как таковой должен представлять широкий интерес для сообщества нейробиологов.
Благодарим за положительный отзыв.
Мы отмечаем, что теперь мы явно изучаем анатомические параметры, которые могут указывать на «сильные стороны отдельных синапсов».Чтобы быть точным, мы описываем вариации в параметрах размера синапса (объем Т-образного стержня, постсинаптическая контактная область). Мы обнаружили, что, хотя эти параметры существенно различаются в разных синапсах, они незначительно различаются в разных синаптических связях, как отмечалось выше.
Рецензент № 3:
Ссылки даются на текстовые страницы, а не на pdf.
Варианты подключения, которые позволяют и ограничивают нейронные вычисления в сенсорной микросхеме.
Во-первых, снимаю шляпу перед авторами за их самоотверженное исследование схем AL с использованием таких трудоемких методов.Для этого представления анатомия является краеугольным камнем, в то время как аспекты моделирования более предположительны. В частности, я бы сказал, что самое слабое звено — это основная сюжетная линия, связь между синаптической структурой и синаптической силой. Даже в этом случае должна быть возможность связать все параметры коннектома с параметрами моделирования, при условии, что у нас есть все необходимые меры. С этой целью авт. Предоставляют важные данные о размере дендритов, которые, как правило, не рассматривались широко в литературе по нейронам дрозофилы.Хотя подсчитать количество синапсов и графики распределения легко, однако в рассказах авторов пока отсутствует размер синапсов. Различается ли это каким-либо согласованным образом между (пре) синаптическими сайтами, и изменяются ли постсинаптические плотности, которые, как я полагаю, также видимы, с размером пресинаптического контакта? Размер дендрита может зависеть от них, но сам по себе не является показателем размера синапса. Если авторы заявляют о взаимосвязи между количеством синапсов и силой синаптической передачи, я думаю, что эти дополнительные параметры должны быть включены, и надеюсь, что их можно относительно легко добыть из существующих данных изображения, даже если я предполагаю, что это все фиксированные модули. .
По просьбе рецензента мы теперь измерили параметры размера синапса. Мы предоставляем обзор нашего анализа и наших выводов в начале этого ответного документа, где мы отвечаем на Резюме редактора (в котором выделен этот конкретный запрос). Анализ размера синапсов теперь включен в рисунок 1 — приложение к рисунку 6.
Введение, четвертый абзац. Дополнительное важное обсуждение точности разводки в мозге можно найти в Takemura et al. (PNAS 112 (Rall, 1964): 13711-13716, 2015), на которые авторы ссылаются, но только позже в подтверждение отсутствия клеток в столбцах продолговатого мозга.Документ PNAS включает ссылки на сравнения между левым и правым гомологами у C. elegans и сравнения между числами синапсов в соседних столбцах мозгового вещества у Drosophila, оба из которых имеют отношение к данным авторов.
Выводы авторов относительно систематических и несистематических вариаций проводников (Введение, последний абзац и далее) отличаются от выводов, полученных из зрительной системы мух (Takemura et al., 2015). Фактически, анализ мозгового вещества указывает на высокий уровень синаптической и проводной точности, с которой дрозофила также способна строить синаптические цепи в своем мозге, в данном случае в зрительной системе.Точность или инвариантность в последнем случае, по-видимому, обусловлена необходимостью надежной передачи на высоких временных частотах, требуемых для зрения, — требование, которое может быть менее требовательным для обонятельного пути.
Мы согласны с тем, что Takemura et al. (2015) следует цитировать более заметно. Мы посвятили этой статье место в Обсуждении, но не во Введении. Теперь мы цитируем эту статью во Введении как пример изменчивости нейронных связей.
Рецензент предполагает, что требования визуальной обработки требуют большей точности нейронной архитектуры, чем требования обонятельной обработки. Мы не думаем, что относительные требования к визуальным и обонятельным вычислениям полностью ясны на данный момент, и мы не согласны с идеей, что обонятельная обработка нетребовательна. Запахи могут колебаться с высокой временной частотой в естественных турбулентных условиях с продолжительностью попадания шлейфа менее 20 мс. Эти временные функции полезны для навигации.Обонятельная трансдукция у Drosophila достаточно быстрая, чтобы отслеживать колебания до 10 Гц. Таким образом, обоняние не намного медленнее, чем зрение. Обоняние также имеет очень большие размеры, потому что для описания различий между различными химическими веществами необходимо большое количество физиолохимических измерений. Это еще одна причина, по которой для обонятельной системы может быть адаптирована высокая скорость передачи информации на периферии. Поэтому особенно интересно, что обонятельная система проводки должна быть неточной.
Мы не думаем, что уровень вариации, который мы здесь описываем, заметно превышает неточность, описанную Takemura et al. (2015). Takemura et al. описал два вида вариаций:
1) Вариация двоичного соединения (отсутствующие соединения или соединения, которые обычно отсутствуют, но иногда присутствуют). В зрительной доле Takemura et al. обнаружили, что скорость двоичной вариации составляет около 1%. В синапсах ORN-PN мы обнаружили, что частота составляет около 2%. (Один ORN слева не смог установить соединение с двумя правыми PN, а один ORN справа не смог установить соединение с тремя левыми PN.Таким образом, частота пропущенных соединений составляет 2 + 3, разделенные на ((26 ORN + 27 ORN) × 5 PN), или 2%. Таким образом, вариации двоичного соединения не сильно различаются в двух наборах данных.
2) Вариация аналогового соединения (изменение количества синапсов на одно соединение для всех экземпляров конкретного типа соединения). В зрительной доле Takemura et al. обнаружили, что CV числа синапсов на одно соединение колеблется от 0,08 до 0,87, в зависимости от типа соединения. В синапсах ORN-PN мы обнаруживаем, что CV равен 0.31. Таким образом, вариации аналогового подключения не сильно различаются в двух наборах данных.
Учитывая изменчивость, описанную Takemura et al., Неудивительно, что мы также находим изменчивость в нашей реконструкции. Вклад нашего исследования не просто в описание изменчивости как таковой. Скорее, вклад этого исследования состоит в том, чтобы предоставить доказательства как систематических, так и бессистемных видов анатомической изменчивости. Нам не нужно предполагать, что вся изменчивость — это «шум» (или «ошибка»).Мы показываем, что некоторые анатомические параметры изменяются систематическим образом, что должно быть адаптивным (учитывая структуру нашей модели), тогда как другие параметры изменяются бессистемным образом, что должно быть неадаптивным (опять же, учитывая структуру нашей модели). .
Прямая связь. Я нахожу «прямую связь» в следующей конструкции ненужной и сбивающей с толку: «вместе со всеми возбуждающими синапсами прямой связи на эти клетки». (Введение, четвертый абзац). «Прямая связь» и «прямая связь» кажутся мне противоречивыми, потому что прямая связь может относиться к синаптическим выходам рассматриваемой клетки.Я думаю, что авторы хотят сказать «вместе со всеми возбуждающими входами (синапсами) в эти клетки».
Мы благодарим рецензента за указание на это. Наше намерение состояло в том, чтобы просто отличить синапсы ORN → PN от других типов возбуждающих синапсов, которые не являются «прямой связью» (например, PN → PN синапсы, PN → ORN синапсы). В отредактированной рукописи эту фразу изменили, чтобы она читалась следующим образом: «вместе со всеми возбуждающими синапсами прямой связи, которые получают эти клетки».
«Три эксперта независимо идентифицировали клубочки DM6 в наборе данных ЭМ путем визуального осмотра на основе опубликованных карт уровня освещенности».Отчет примечателен скорее тем, что не говорит, чем тем, что он говорит. Я считаю, что документации о том, как авторы выбрали нейроны DM6 для реконструкции, недостаточно. Хотя конфокальные данные ясно показывают клубочковую карту антеннальной доли, особенно на основе синаптической и генетической маркировки, такие данные вводят в заблуждение для электромагнитных исследований, которые выявляют все профили, а не их селективные особенности. В частности, нет четких глиальных границ между клубочками и клубочками, о которых авторы не упоминают, и которые могут быть однозначно идентифицированы только со ссылкой на карту окружающих клубочков или, возможно, по картезианским координатам, и я думаю, что авторы должны это признать. .Вполне возможно, что авторы приобрели достаточный опыт из физиологических записей, чтобы идентифицировать специфические клубочки, но это находится в интактной живой доле усиков. Клубочки также частично перекрывают друг друга в глубине нейропиле доли. Более того, нам не сообщают, почему сам DM6 был выбран по сравнению с другими клубочками, и в качестве обоснования авторы могут захотеть процитировать свои собственные физиологические исследования. Эти вопросы являются центральными для анализа авторов и должны быть разъяснены в тексте, потому что, хотя я уверен, что были предприняты большие усилия, и мы можем быть уверены, что три эксперта сделали одну и ту же идентификацию, на самом деле это нетривиальная проблема. все на уровне EM.Большинство читателей не знают или не понимают этого, потому что конфокальные карты настолько убедительны, потому что большинство читателей никогда не видят ЭМ и потому что большинство может ошибочно предположить, что существует четкая глиальная граница, окружающая каждый клубочек. Также вполне возможно нацелить DM6 на живой препарат, что неоднократно проводилось в исследованиях лаборатории Уилсона, но контуры поверхности антенны также отличаются от 2D-изображений из ЭМ-срезов. Эти вопросы «почему» и «как» необходимо четко решить.Текст Методики должен позволять другому работнику повторить наблюдения авторов. В частности, как авторы могут предоставить читателю четкую уверенность в том, что они на самом деле нацелены на одни и те же клубочки на левом и правом полушариях мозга, и что левая / правая асимметрия, которую они обнаруживают, не являются результатом выбора разных клубочков на этих двух? стороны. Напротив, я полностью согласен с тем, что авторы скрупулезно идентифицируют в наборе данных EM ORN и PN из их соответствующих ветвей и траекторий аксонов.
Границы клубочков антенных долей очерчены специальным классом глии (называемым «обволакивающей глией»; Doherty et al. 2009 J Neurosci29: 4768-81). Мы согласны с рецензентом в том, что эти глиальные оболочки могут быть более или менее четкими в материале ssEM, в зависимости от того, как был подготовлен образец и визуализирован, а также в зависимости от идентичности рассматриваемых клубочков. В нашем материале границы многих клубочков довольно четкие при низком разрешении (см. Видео 1-2).Мы решили сосредоточиться на DM6 клубочка, отчасти потому, что это выступающий клубок, с легко определяемым положением относительно клубочков других долей усиков. На опубликованных картах антеннальной доли DM6 имеет особенно последовательное расположение (Laissue et al., 1999 J Comp Neurol405, 543-552; Couto et al. 2005 Curr Biol15, 1535-1547; Grabe et al. 2015 J Comp Neurol523, 530 -544). Мы также выбрали DM6, потому что имеется большой объем физиологической информации об этом клубочке. Наконец, DM6 был расположен в области серии EM, где имеется относительно мало недостающих секций (подробно описано в исходных материалах и методах).Теперь мы пересмотрели методы, чтобы более четко объяснить наше обоснование выбора DM6. Тот факт, что мы правильно идентифицировали пару гомологичных клубочков на двух сторонах мозга, был в конечном итоге подтвержден, когда была проведена реконструкция: мы обнаружили, что аксоны ORN DM6 пересекали переднюю комиссуру, чтобы нацеливаться на другой клубок, который мы идентифицировали как DM6.
Рисунок 1A. Связанный: хотя и полезен как обзор EM, рисунок 1A не дает достаточной информации для идентификации DM6, особенно по оси z.Был ли реконструирован весь клубочек и как авторы определили, где лежат его границы?
Благодарим рецензента за возможность прояснить этот момент. Некоторые границы DM6 (а именно передняя, медиальная и дорсальная границы) были четкими при визуальном осмотре (см. Видео 1-2). Задняя, латеральная и вентральная границы были менее четкими, и поэтому в конечном итоге определялись дендритами реконструированных PN. Все дендриты реконструированных нами PN заполнены полностью одинаковым объемом (что типично для большинства PN и большинства клубочков).Как описано в Методах, мы реконструировали каждый профиль пресинаптического профиля для каждого дендрита PN (по крайней мере, до границы DM6), и поэтому мы с высокой степенью уверенности идентифицировали каждый ORN DM6. Цель рисунка 1A — предоставить обзор ЭМ-разреза на уровне DM6, но идентификация DM6, конечно же, была сделана с помощью 3D-данных с высоким разрешением. Мы ожидаем, что специализированные читатели, заинтересованные в этом выпуске, изучат набор трехмерных данных на сайте neurodata.io.
Сироты: К чему относится процент сирот? Число независимых фрагментов, которые были пресинаптическими по отношению к PN, но не могли быть связаны с идентифицированной клеткой? Сколько синапсов они внесли? Были ли осиротевшие с обеих сторон, не имевшие контакта ни с идентифицированным ORN, ни с PN? Если да, то в какой пропорции?
Мы определяем сирот как фрагменты реконструированных нейритов, которые не могут быть связаны ни с одним нейроном.Теперь мы изменили текст, чтобы уточнить это определение. Автор обзора прав, что сироты были независимыми фрагментами, которые были пресинаптическими по отношению к PN, но не могли быть связаны с идентифицированной клеткой. С нашей стратегией реконструкции (реконструировать все входы в PN) один конец всегда привязан к входному синапсу PN, поэтому мы не генерируем фрагменты, потерянные на обоих концах. Следовательно, по конструкции количество синапсов, внесенных сиротами в PNs, по существу является количеством самих сиротских фрагментов.
Утверждение (подраздел «Клетки, составляющие клубочковую микросхему», последний абзац) о том, что ложные продолжения между фрагментами легко обнаруживаются и исправляются во время независимого обзора, скорее является признаком ветки, чем стратегии реконструкции, поскольку личинка нейроны у Drosophila, как правило, имеют относительно более прочные нейриты с дистальными конечностями, несущими большинство синапсов. Таким образом, вывод о том, что они очень редки при окончательной реконструкции, может быть менее обоснованным, чем утверждается.
Хотя ложные продолжения возможны, наша стратегия реконструкции способствует минимизации ложных продолжений с риском ложных завершений, чтобы максимизировать уверенность в восстанавливаемых нами соединениях. Аннотаторы перестают отслеживать
, если продолжение становится неоднозначным. Более того, все реконструированные нейроны были просмотрены несколькими независимыми аннотаторами.
Пресинаптические Т-образные стержни. Прокомментируйте, пожалуйста, диапазон структурных фенотипов, демонстрируемых пресинаптическими Т-образными полосками на Рисунке 1 — приложение к рисунку 3, или из анализа авторов синапсов в последовательных срезах.
Теперь мы включили количественный анализ объемов Т-образных стержней в исправленную рукопись (Рисунок 1 — приложение к рисунку 6), как описано выше. Мы считаем, что подробное описание форм Т-образных стержней выходит за рамки данного исследования, но мы планируем изучить этот вопрос в будущем.
Таблица 1. Подраздел «Относительное количество типов подключения», первый абзац. Я не нашел Таблицу 1, которая необходима для интерпретации Рисунка 1 — дополнения к рисунку 4.
Рецензент, возможно, не смог найти Таблицу 1, поскольку она представлена в виде отдельного машиночитаемого дополнительного файла со значениями, разделенными запятыми (CSV).
Синаптические числа, подраздел «Структура и сила возбуждающих связей». Документирование номеров синапсов хорошее, и интерпретация, как правило, уместна, но авторы должны внимательно просмотреть свой текст, чтобы проверить тенденцию приравнивать количество синапсов к силе синапсов и строго различать семантические ссылки на эти два.
Мы явно отмечаем во введении, что сила синаптической связи может зависеть от многих анатомических особенностей.Количество синапсов на одно соединение — лишь одна из этих функций. Одна из целей нашего исследования состояла в том, чтобы сделать вывод, какие особенности на самом деле вносят наибольший вклад в наблюдаемые физиологические вариации силы связи. Мы начинаем с того факта, что ипсилатеральные связи ORN → PN, как известно, на 30-40% сильнее, чем контралатеральные связи (Gaudry et al. 2013), и мы спрашиваем, какие анатомические особенности могут объяснить это физиологическое различие. Мы обнаружили, что количество синапсов на соединение примерно на 27% выше на ипсилатеральной стороне (рис. 4D), в то время как нет систематической разницы в дендритных свойствах ипсилатеральных и контралатеральных синапсов (судя по амплитудам mEPSP, которые отражают расположение синапсов на дендрите). и электрические фильтрующие свойства самого дендрита, рис. 4E).Этот результат свидетельствует о том, что количество синапсов является основным фактором изменения прочности соединения. Более того, мы не обнаружили существенной разницы между ипси- и контралатеральной зоной постсинаптического контакта ПП. Контралатеральные Т-образные стержни оказываются на 8% больше, чем ипсилатеральные Т-образные стержни, но эта разница относительно невелика, и это объясняется тем фактом, что Т-образные стержни растут с увеличением количества постсинаптических элементов, а контрлатеральные проекции ORN нацелены на большее количество элементов. профили без PN, чем у ипсилатеральных ORN (Рисунок 4 — приложение к рисунку 1).Наконец, когда мы просто сосредотачиваемся на ипсилатеральных связях, мы снова обнаруживаем, что количество синапсов на одно соединение снова является наиболее изменчивой анатомической характеристикой синаптических связей (CV = 0,31, рис. 5C) со сравнительно меньшими вариациями общих дендритных свойств соединения (CV амплитуда mEPSP = 0,014, Рисунок 5E; Эффективность суммирования CV = 0,041, Рисунок 5G) или средний размер синапса для каждого соединения ( объем CVT-стержня = 0,13, площадь постсинаптического контакта CV = 0.13, рисунок 1 — приложение к рисунку 6г). Взятые вместе, эти результаты свидетельствуют о том, что число синапсов является основным анатомическим признаком, влияющим на вариации прочности соединения, хотя другие анатомические особенности также вносят свой вклад. Это важный результат для данной области, потому что он устанавливает номер синапса в качестве разумного показателя прочности соединения — с оговоркой, что эта функция определенно не улавливает все изменения в силе соединения.
В контексте рукописи мы используем фразу «сила связи» только тогда, когда намеренно говорим о силе физиологической связи — либо в контексте опубликованных исследований, в которых непосредственно измерялись синаптические реакции, либо в контексте нашей компартментальной модели, который использует параметры, измеренные в опубликованных электрофизиологических исследованиях, для экстраполяции наших анатомических измерений на физиологические прогнозы.Мы подчеркиваем, что число синапсов является разумным показателем силы соединения, но мы также поясняем, что это всего лишь приблизительный показатель, потому что число синапсов — не единственная анатомическая характеристика, которая вносит вклад в вариации между синаптическими связями.
Также важно помнить о глубине обработки, вносимой синапсами обратной связи, для которых усиление передачи отражает не только количество синапсов прямого распространения, но и синапсов восходящего направления. Таким образом, помимо усиления отдельных синапсов, схемы обратной связи наследуют усиление сигнала со своих синаптических входов.
Мы согласны с рецензентом в том, что соединения с прямой связью, на которых мы фокусируемся в этом исследовании (соединения ORN → PN), не являются единственными важными элементами этой схемы. Мы действительно сообщаем данные о PN → PN синапсах, ORN → ORN синапсах и PN → ORN синапсах, хотя мы не фокусируемся ни на одном из этих типов соединений в последней части исследования. Мы также четко заявляем о том, что в цепи есть много клеток, которые мы не полностью реконструировали (клетки, которые мы называем мультигломерулярными клетками, которые в основном являются локальными нейронами).Конечно, некоторые из этих дополнительных клеток, вероятно, будут опосредовать обратную связь. Мы ожидаем, что элементы обратной связи этой схемы (например, локальные нейроны, которые опосредуют подавление обратной связи) будут описаны в будущих исследованиях ssEM.
Матрица смежности на рис. 1 — дополнение к рисунку 4 напоминает ряд таких матриц, которые были опубликованы, с верхними числами около 50 синапсов, что соответствует таковым в других областях мозга, на которые авторы могли указать. Каждый перехват представляет собой место предполагаемой передачи между одним пре- и постсинаптическим нейроном, но в тексте следует признать, что, поскольку каждый пресинаптический сайт имеет несколько постсинаптических элементов, ряд перехватов в матрице обязательно скоординированно связаны.Фактически, я не нахожу простой метрики синаптической дивергенции, показанной на этом рисунке или где-либо еще в тексте. Сколько диад, триад и т. Д. Включено в матрицу. Как вариант, сколько монад?
Мы согласны с рецензентом, что эта информация должна быть сообщена. Мы добавили рисунок 1 — приложение 5, показывающее распределение полиад в выходных синапсах ORN в DM6.
Относится к предыдущему пункту: Рисунок 1 — дополнение к рисунку 3. Это отдельные изображения синапсов, которые не могут выявить профили выше и ниже плоскости сечения.Тем не менее, очевидно, что на сайтах единственного выпуска есть несколько дендритов, что действительно можно увидеть в некоторых разделах. Эта информация представлена на Рисунке 4 — приложение к рисунку 1E, но только в виде средних значений для сравнения ORN Ipsi и Contra.
См. Ответ на предыдущий пункт рецензента. Теперь мы более четко проясним существование диад, триад и т. Д. На Рисунке 1 — добавлении к рисунку 5.
Авторы заявляют, что «Все события синаптической проводимости имели одинаковый размер и форму.«Это предположительно предполагает, что эти события проводимости были общими в одном и том же пресинаптическом участке? Представляют ли авторы, что каждый постсинаптический дендрит видит одно и то же облако высвобождения нейротрансмиттера, или это облако разделяется между несколькими дендритами. Если это число варьируется, означает ли это, что эта передача зависит от количества дендритов, или дендритный сигнал не изменяется; как результат влияет на моделирование авторов? Я не думаю, что есть простой ответ, но эти проблемы следует, по крайней мере, признать, и насколько возможно обсуждается.
Когда мы говорим: «Все события синаптической проводимости имели одинаковый размер и форму», мы имеем в виду все события проводимости на всех постсинаптических участках периода. По сути, наша модель предполагает, что появление спайка на Т-образной полосе всегда открывает одинаковое количество постсинаптических ионных каналов во всех клетках, постсинаптических по отношению к этой Т-образной полосе. Мы изменили текст рукописи, чтобы сделать это предположение как можно более четким. Это предположение, очевидно, является упрощением, но построение модели всегда включает в себя некоторые упрощения, и мы думаем, что это не является необоснованным упрощением.Конечно, важно, чтобы одни Т-образные стержни были привязаны к одному постсинаптическому профилю, а другие — к двум профилям (диадам) или трем профилям (триадам) или даже большему количеству. Однако мы не думаем, что синаптические проводимости обязательно слабее в случаях, когда большее количество дендритов «разделяют» одну и ту же Т-образную перемычку, потому что нет причин, по которым концентрации нейротрансмиттеров обязательно ниже в диадических или триадных связях по сравнению с монадическими связями. Примечательно, что мы находим, что объем Т-образного стержня растет с увеличением количества профилей, постсинаптических по отношению к Т-образному стержню (Рисунок 1 — приложение к рисунку 6А), что может указывать на механизм, посредством которого квантовое содержание увеличивается, чтобы соответствовать объему синаптической щели.
Рисунок 1 — дополнение к рисунку 4. Кроме того, я считаю, что использование черного цвета для 0 синапсов сбивает с толку, не в последнюю очередь потому, что мне трудно различать различия в тоне для темных перехватов с небольшим количеством синапсов, и я рекомендую перестроить эту матрицу, используя белый цвет для перехватывает 0 синапсов. Как вариант, переверните весь спектр тонов, чтобы 50+ был черным, а 0 — белым.
Благодарим рецензента за это предложение. Пожалуйста, ознакомьтесь с рисунком 1 — дополнением к рисунку 4, обновленным в новой редакции.
Методы. В разделе «Крупномасштабная визуализация и совмещение изображений с помощью ПЭМ» нам сообщают, сколько секций было потеряно, по всей видимости, 145, от 1 до 6, около 7% от общей серии — примерно столько же количество сирот, но не количество срезов, необходимых для охвата клубочков DM6, и, следовательно, не то, какая часть из них была каплями. Потеря 6 секций (0,3 мкм) наносит большой ущерб непрерывности любой серии, но, по крайней мере, авторы честно сообщают о них.Интересно, сколько из этих капель привело к появлению сирот, и были ли они равны на левой и правой сторонах DN6? Авторы, я думаю, должны попытаться решить эти вопросы.
В пересмотренных Методиках мы приводим следующие детали относительно утраченных секций: «В пределах части объема, охваченной клубочками DM6, было 9 одиночных потерь, 2 случая 2-сегментных потерь, 1 случай 3-сегментных потерь. , и 2 случая 4-х секционных потерь (потери относятся к последовательным секциям).Конечно, потерянные разделы прискорбны, но они также не редкость в такого рода наборах данных. Мы выбрали DM6 специально, потому что ни один из 5-секционных или 6-секционных потерь не пересекал этот клубок. Некоторые из бесхозных фрагментов, которые мы реконструировали, являются следствием потери сечения, но другие бесхозные фрагменты не примыкают к потерям сечения, а вместо этого возникают из-за неоднозначности, связанной с тем, как плоскость среза пересекает нейрит.
https://doi.org/10.7554/eLife.24838.021(PDF) Электрические испытания многосерийных образцов микросхем малой серии, сочетающих технологии Wire Bond и Flip-Chip
Электрические испытания многосерийных многокристальных микросхем
образцов, сочетающих технологии Wire Bond и Flip-Chip
Д.А. Кириенко Ершова Н.Ю., Путролайнен В.В., Лунков П.В., КР
Марцинкевич
Физико-технический институт Петрозаводского государственного университета, ул. Ленина, 33,
185910, Петрозаводск, Россия,
E-mail: kirienko @ petrsu.ru
Аннотация. В статье представлено техническое решение и методика проведения автоматизированных электрических и функциональных испытаний
малых серий многочиповых сборок. Представлены результаты испытаний
образцов многокристальных сборок, сочетающих технологии Wire Bond и Flip-Chip. По результатам испытаний локализованы дефекты, возникающие в процессе упаковки,
и определены этапы сборки, на которых эти дефекты возникают.
1. Введение
Основным направлением развития современной микроэлектроники является постоянное уменьшение массы изделий и габаритов
, повышение их производительности и надежности. В связи с этим перспективным является использование электронной компонентной базы
, интегрированной в систему в корпусе (SiP) [1, 2]. SiP представляет собой набор кристаллов процессора
, памяти, блоков цифровой обработки сигналов или других компонентов, соединенных между собой и расположенных в едином корпусе
на подложке.Кристаллы расположены на одном уровне или один над другим и
дополнены пассивными или другими необходимыми компонентами. Несколько подложек с кристаллами
могут быть уложены друг на друга по технологии Package on Package (PoP) с припаиванием шариковых выводов на
верхних подложек к контактным площадкам на нижних подложках [3]. Как правило, для монтажа кристаллов используется технология Flip-Chip или Wire
Bond. Кристалл Flip-Chip устанавливается на плату активной стороной вниз
и фиксируется с помощью шаровидных контактов.В случае технологии Wire Bond кристалл
устанавливается непосредственно на подложку, а проволочные контакты прикрепляются к подложке или другому элементу путем сварки
[4]. Каждый микроэлектронный компонент SiP имеет одно или несколько независимых целей. Кроме того, каждый компонент
может быть частью других элементов в SiP. Важным преимуществом парадигмы SiP является
, что все элементы системы могут быть изготовлены отдельно, с использованием разных технологий, а затем
собраны в одно конечное электронное устройство [5].Такой подход выгоден как производителям микросхем,
, позволяющих крупномасштабное производство микроэлектронных компонентов, так и их клиентам, предоставляя
возможность быстро и легко разрабатывать специализированные электронные устройства на основе этих компонентов [5].
Одним из важных этапов разработки новых интегральных схем (ИС) на основе технологий SiP / PoP
является прототипирование. При прототипировании создается макет разрабатываемого изделия
с частичной или полной заменой основных функциональных компонентов (кристаллов) на макетные микросхемы
с такими же размерами и количеством выходов [6, 7].На прототипе отрегулирована вся производственная линия
, включая тестирование интегральных схем, этапы маркировки, сканирования и упаковки.
В процессе производства интегральной схемы в электрической части изделия могут возникать различные дефекты, влияющие на ее функциональность. Например, на этапе пайки кристалла Flip-Chip
могут образоваться дефекты, такие как перемычки между соседними шариковыми выводами, непаянные соединения кристалла с кристаллом
НАПЕЧАТАНО ИЗ ОНЛАЙН-СТИПЕНДИИ ОКСФОРДА (Оксфорд.Universitypressscholarship.com). (c) Авторские права Oxford University Press, 2021. Все права защищены. Отдельный пользователь может распечатать одну главу монографии в формате PDF в OSO для личного использования. дата: 15 июля 2021 г.
- Глава:
- (стр.195) 10 заключительных мыслей: структура теории кортикальной микросхемы
- Источник:
- Разнообразие в нейронной машине
- Автор (ы):
Иван Солтес
- Издатель:
- Oxford University Press
В этой главе представлен синтез обсуждений в предыдущих главах. Он утверждает, что крайне важно четко различать гетерогенность, изменчивость и разнообразие, поскольку большая часть разногласий в литературе возникла из-за смешения аргументов, связанных с межклеточной изменчивостью в рамках определенных межнейронных подтипов, с данными о межнейрональном видовом разнообразии. Межклеточная изменчивость не только естественным образом существовала в популяциях интернейронов, принадлежащих к одному подтипу, но и то, что эта межклеточная изменчивость имела несколько функциональных последствий, включая регуляцию возбудимости нейронов, осцилляторную активность и сетевую синхронность.
Ключевые слова: интернейроны, межнейронное разнообразие, шанс, нейроанатомия
Для получения доступа к полному тексту книг в рамках службы для получения стипендииOxford Online требуется подписка или покупка. Однако публичные пользователи могут свободно искать на сайте и просматривать аннотации и ключевые слова для каждой книги и главы.
Пожалуйста, подпишитесь или войдите, чтобы получить доступ к полному тексту.
Если вы считаете, что у вас должен быть доступ к этой книге, обратитесь к своему библиотекарю.
Для устранения неполадок, пожалуйста, проверьте наш FAQs , и если вы не можете найти там ответ, пожалуйста свяжитесь с нами .
ИСПЫТАНИЕ МИКРОСХЕМ ВОЕННОГО КЛАССА — Gemes
MIL-STD-883 — это стандарт Министерства обороны США для методов тестирования микросхем. На более чем 700 страницах он охватывает широкий спектр требований к испытаниям, от требований к калибровке и температуре до деталей методов внутреннего контроля микросхем.Помимо общих требований, существует более 100 конкретных методов испытаний. Последнюю редакцию (Rev. K с изменением 2) полного документа можно загрузить в Defense Logistics Agency по адресу: https://landandmaritimeapps.dla.mil/Programs/MilSpec/listdocs.aspx?BasicDoc=MIL-STD-883 .
МЕТОДЫ ИСПЫТАНИЙ MIL-STD-883
Различные методы испытаний в стандарте MIL-STD-883 разделены на 4 класса. Каждому классу присваивается свой диапазон номеров:
1001-1999 — Экологические испытания
2001-2999 — Механические испытания
3001-4999 — Электрические испытания
5001-5999 — Процедуры испытаний
Эти методы испытаний определяют оборудование, настройку и методы измерения, которые будут использоваться.Поскольку методы испытаний применяются к широкому кругу различных частей, многие методы испытаний указывают на то, что подробные сведения, относящиеся к конкретному устройству, должны быть включены в подробную спецификацию для этой части. Например, метод 3007 испытания для низкого уровня выходного напряжения указывает, что температура испытания, ток принуждения, напряжение (я) источника питания, входные уровни и пределы «должны быть указаны в применимом документе о приобретении». Поскольку эти методы тестирования предназначены для охвата всего диапазона микросхем, не все тесты применимы к каждому устройству.Документ о приобретении (а / к / спецификация на закупку) должен указывать, какие тесты требуются для этой части, а также детали, необходимые для различных стандартных тестов.
НАШИ МИКРОСХЕМЫ КЛАССА B СООТВЕТСТВУЮТ ВЫСОКОЙ НАДЕЖНОСТИ ВОЕННЫХ ПРИЛОЖЕНИЙ
Однако методы в разделе «Процедуры тестирования» несколько отличаются. В частности, два из этих методов требуют выполнения целых групп других стандартных тестов: метод 5004 «Процедуры проверки» и метод 5005 «Требования к квалификации и качеству».Метод 5004 тестирования определяет различные этапы проверки, которые должны выполняться на каждой микросхеме военного качества. Эти шаги могут различаться в зависимости от уровня сертифицированного класса качества: класс B — обычное высоконадежное военное приложение; класс S — приложение космического уровня. Устройства GEM проходят проверку и сертифицируются на уровень класса качества B. Для уровня класса качества B на протяжении всего производственного процесса требуется несколько экранов: приемочные испытания партии вафель; внутренний визуальный осмотр; температурный цикл; постоянное ускорение; внешний визуальный осмотр; высокотемпературное пригорание; Расчет допустимого процента брака (КПК); Заключительное электрическое испытание при высоких, низких и комнатных температурах; испытание герметичности упаковки.После того, как производственная партия деталей соответствует требованиям проверки 5004, она подлежит проверке качества (QCI) по методу 5005. Для этого требования к испытаниям группы образцов выбираются случайным образом из производственной партии для «Групповых испытаний». Для деталей класса качества B без радиационной гарантии существует 4 группы: Группа A (электрические испытания) — 116 образцов, Группа B (разрушающие механические испытания и испытания на воздействие окружающей среды) — 3 образца, Группа C (испытание на долговечность) — 45 образцов, Группа D (тест, связанный с пакетом) — размер выборки варьируется.