
Что такое коробка уравнивания потенциалов и как её подключают к СУП?
В соответствии с современными нормами, в здании, где проложена электропроводка, должна иметься система уравнивания потенциалов. Причем, помимо основной, устанавливается и дополнительная СУП. Об организации последней (подключение коробки уравнивания потенциалов и т.д.) пойдет речь в этой статье. Начнем с теории, а именно, что представляют собой СУП.
Пример организации основной и дополнительной СУПНазначение
Металлические конструкции здания, проложенные в нем инженерные и коммуникационные сети, являются проводниками электричества. Пока они не замкнуты в единый контур между этими элементами может возникнуть разность потенциалов. Причем, причиной этому может быть как повреждение изоляции силового кабеля, так и статика, наводка, атмосферное перенапряжение и т.д. Это грозит тем, что коснувшись, например, корпуса бытового электроприбора и радиатора отопительного контура можно получить не только ощутимый, а и фатальный удар током.
Чтобы избежать таких неприятностей, все токопроводящие конструкции в здании, а также инженерные сети с подобными свойствами, объединяют в единый контур, соединенный с клеммной колодкой PE. Заземление может быть проложено отдельно или подводиться совместно с входящим кабелем.
При правильно организованной СУП в случае «пробоя» на токопроводящие элементы конструкции произойдет короткое замыкание или возникнет большой ток утечки, что приведет к активации устройств, производящих отключение. В результате поврежденный участок будет отключен и угрозы человеческой жизни не возникнет.
Система уравнивания потенциалов обеспечивает защиту в случае пробояОсновная СУП
Она закладывается, непосредственно, при строительстве здания либо в процессе его реконструкции или капитального ремонта.
Основные составляющие СУП:
- Заземляющий контур.
- Провода РЕ.
- Главная заземляющая шина.
Не будем вдаваться в подробности организации основной СУП, поскольку это тема отдельной статьи, но приведем три догмата ее построения:
- Объединять провода, используемые СУП с рабочим нолем (N), категорически запрещается.
- Заземление должно проводиться по радиальной схеме, то есть к каждому элементу подводиться отдельный проводник.
- В системе нельзя устанавливать никакие коммутационные устройства, поскольку одно из основных требований гласит, что входящие в нее проводники должны быть беспрерывны.


Необходимость дополнительной СУП и ее основные элементы
Казалось бы, зачем устанавливать дополнительную систему, если есть основная, на это есть несколько весомых причин, приведем две из них:
- Обеспечение безопасности во влажных помещениях, где установлено электрооборудование (например, ванная комната и кухня).
- Нет гарантии, что сосед снизу в процессе ремонта не заменил в стояке или контуре отопления металлические трубы на пластиковые, нарушив тем самым основную СУП.
Как видите, этих двух причин вполне достаточно для возникновения потенциальной угрозы жизни.
Основные элементы дополнительной системы:
- Коробка уравнивания потенциалов (КУП), снабженная специальной клеммной колодкой (ШДУП) для дополнительного подключения элементов с токопроводящими свойствами. Желательно выбрать изделие известных брендов, например, DKC, Hegel или отечественные модели КУП2604, КУП2603, КУП1101 и т.д. Размер коробки подбирается в зависимости от количества подключаемых элементов.
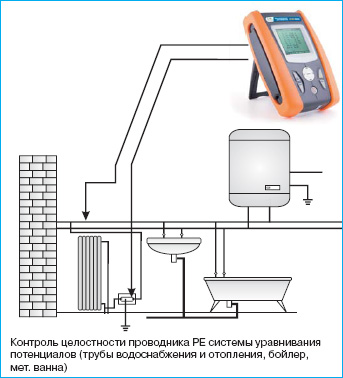
В принципе, можно использовать и коробки неизвестных брендов из Поднебесной, только прежде, чем производить установку, убедитесь, что для их изготовления использовался негорючий материал. Помимо этого проверьте качество исполнения внутренней клеммной колодки. Было немало случаев, когда крепление не обеспечивало надежного контакта.
- Провода, используемые ДСУП. Их количество должно соответствовать числу подключаемых элементов. За основу берется медный провод, у которого сечение в пределах 2,5-6 мм. Сразу заметим, что алюминиевые жилы не подходят для этой задачи, поскольку образующаяся на нем оксидная пленка может нарушить контакт, сведя на нет функциональное назначение системы.
Организация дополнительной системы
Весь процесс можно условно разделить на два этапа подготовительный и, собственно, монтаж.
В первую очередь необходимо составить схему ДСУП, хотя бы приблизительную. Это позволит «не намудрить» с подключениями, например, забыть подсоединить к клеммной колодке какой-либо элемент или подвести к нему несколько проводов РЕ. Пример правильно выполненного эскиза показан на рисунке 5.
На эскизе показаны элементы, которые следует в обязательном порядке подключить к КУП, перечислим их:
- 1 и 2 – трубы подачи холодной и горячей воды.
- 3 – труба канализационной системы.
- 4 – корпус ванны (если она металлическая, как вы понимаете, акриловую ванну заземлять не имеет смысла).
- 5 – радиатор отопления (если таковой имеется в ванной комнате).
- 6 – экран системы «теплый пол».
- 7 – дверной короб (если он из металла).
- 8 – вентиляционная решетка (при условии, что в системе вентиляции используются металлические трубы).
При правильно организованной электропроводке подводить отдельно провода к электроприборам не имеет смысла, поскольку эту функцию выполняет жила заземления.
Сделав эскиз, необходимо определиться, где будет устанавливаться КУП, особых критериев к этому нет, главное, чтобы к ней непроблематично было добраться в любое время.
Определившись с местоположением коробки, следует продумать трасы для проводов, идущих к КУП. Напоминаем, что они должны быть уложены по радиальной схеме, то есть к каждому элементу идет отдельный провод.
Если при подключении проводников системы к коробке с клеммником проблем не будет, то с заземлением труб могут возникнуть некоторые сложности. Исправить ситуацию можно с помощью специальных хомутов зажимов, один из них продемонстрирован на рисунке 6.
Рис 6. Зажим для подключения защитного заземления к трубамМесто для крепления провода заземления к хомуту отмечено красным кругом.
После того, как ДСУП смонтирована, клеммник КУП подключается к РЕ квартирного щитка (см. рис. 7).
Рис 7. Подключение ДСУП к шине РЕ
После установки системы и подключения ее клеммной колодке PE необходимо убедиться в наличии между нею, КУП и подключенными элементами единой цепи. Сделать это можно произведя соответствующие замеры.
В остальных комнатах организация дополнительной системы не имеет смысла.
Что ещё необходимо принять во внимание?
Дополнительную систему запрещается монтировать, если в доме используется заземление по типу TN-С, поскольку это создает реальную опасность для жизни другим жильцам, не установившим ДСУП.
Подключение КУП к клеммам щитка необходимо выполнять, только обесточив электропроводку.
Обратим внимание! Если в ванной комнате или на кухне установлен бойлер, он должен быть обязательно подключен к КИП, а питание на него необходимо подавать через отдельную линию, защищенную УЗО.
Если на установленные в ванной розетки может попасть вода, то необходимо использовать изделия классом не ниже IP54.
Нужна ли ДСУП в санузле?
Наверняка многие из вас знают правильный ответ, но, все-таки хочется обсудить данную тему.
Сразу хотелось бы акцентировать ваше внимание, что речь идет о санузлах, а не душевых и ванных помещениях.
Санузел – помещение, где установлен унитаз и умывальник. Иногда могут еще разместить полотенцесушитель.
Был у меня один проект, который прошел госстройэкспертизу и имеет положительное заключение, где я не предусмотрел ДСУП в санузле. При вводе объекта в эксплуатацию, энергонадзор потребовал выполнить ДСУП в санузле. Решить данный вопрос с энергонадзором не получилось, поэтому пришлось написать официальное письмо, чтобы указали конкретно ссылки на нормативные документы. А пока энергонадзор думает, как обосновать их требование, я в этой статье попытаюсь им помочь.
Нужно отметить, что санузел относится к влажным помещениям [1], [5].
Относится ли помещение санузла к помещениям с повышенной опасностью? На мой взгляд, вопрос спорный. Как правило, в с/у пол из керамической плитки, а проводимость керамической плитки не совсем такая как железобетонного пола.
Как правило, в с/у пол из керамической плитки, а проводимость керамической плитки не совсем такая как железобетонного пола.
Если считать пол из керамической плитки токопроводящим, то у нас все торговые помещения нужно относить к помещениям с повышенной опасностью. Может и там будем делать ДСУП, заведем в каждую розетку дополнительный PE-проводник?
РД 39-22-113-78 (Временные правила защиты от проявлений статического электричества на производственных установках и сооружениях нефтяной и газовой промышленности):2.5.14. Покрытие пола считается электропроводным, если электрическое сопротивление между металлической пластинкой площадью 50 см2, уложенной на пол и прижатой к нему силой 25 кгс, и контуром заземления не превосходит 10 Ом.
Приложение 1. 1.5. Электропроводные материалы — материалы, имеющие удельное объемное электрическое сопротивление не более 105 Ом·м.
Удельное объемное электрическое сопротивление различных покрытий полов
Исходя из этой информации, даже влажный пол из керамической плитки не является электропроводным.
Я нигде не встречал требований, что в помещениях с/у нужно предусматривать дополнительную систему уравнивания потенциалов.
19.19 В ванных и душевых помещениях должна быть выполнена дополнительная система уравнивания потенциалов в соответствии с требованиями ГОСТ 30331.11 [1].4.3.5.9 Система дополнительного уравнивания потенциалов должна соединять между собой все одновременно доступные прикосновению открытые проводящие части стационарного электрооборудования и сторонние проводящие части, включая доступные прикосновению металлические части строительных конструкций здания, а также нулевые защитные проводники в системе TN и защитные заземляющие проводники в системах IT и ТТ, включая защитные проводники штепсельных розеток.
Для уравнивания потенциалов могут быть использованы специально предусмотренные проводники либо открытые и сторонние проводящие части, если они удовлетворяют требованиям 4. 3.14.2 к защитным проводникам в отношении проводимости и непрерывности электрической цепи[2].
3.14.2 к защитным проводникам в отношении проводимости и непрерывности электрической цепи[2].
8.7.20 Для саун, ванных и душевых помещений дополнительная система уравнивания потенциалов является обязательной и должна выполняться по ГОСТ 30331.11 и с учетом следующих требований [2].
1.7.9. Открытая проводящая часть — доступная прикосновению проводящая часть электроустановки, нормально не находящаяся под напряжением, но которая может оказаться под напряжением при повреждении основной изоляции [4].
1.7.10. Сторонняя проводящая часть — проводящая часть, не являющаяся частью электроустановки [4].
7.1.88. К дополнительной системе уравнивания потенциалов должны быть подключены все доступные прикосновению открытые проводящие части стационарных электроустановок, сторонние проводящие части и нулевые защитные проводники всего электрооборудования (в том числе штепсельных розеток).
Для ванных и душевых помещений дополнительная система уравнивания потенциалов является обязательной и должна предусматривать, в том числе, подключение сторонних проводящих частей, выходящих за пределы помещений.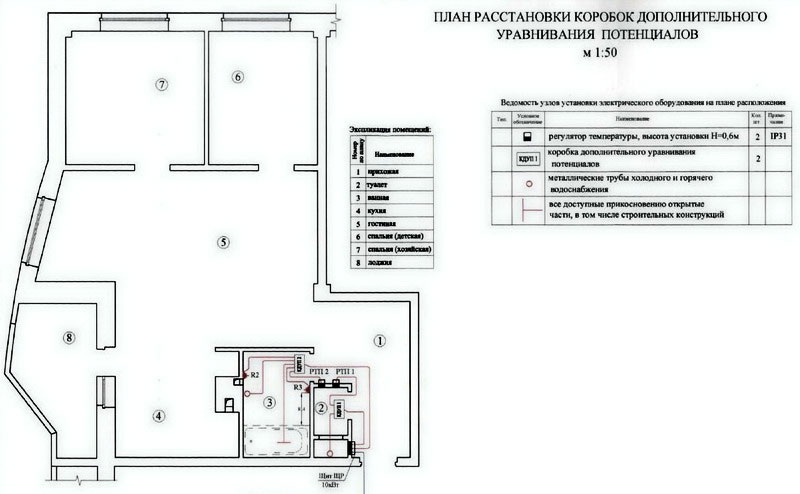 Если отсутствует электрооборудование с подключенными к системе уравнивания потенциалов нулевыми защитными проводниками, то систему уравнивания потенциалов следует подключить к РЕ шине (зажиму) на вводе. Нагревательные элементы, замоноличенные в пол, должны быть покрыты заземленной металлической сеткой или заземленной металлической оболочкой, подсоединенными к системе уравнивания потенциалов. В качестве дополнительной защиты для нагревательных элементов рекомендуется использовать УЗО на ток до 30 мА.
Если отсутствует электрооборудование с подключенными к системе уравнивания потенциалов нулевыми защитными проводниками, то систему уравнивания потенциалов следует подключить к РЕ шине (зажиму) на вводе. Нагревательные элементы, замоноличенные в пол, должны быть покрыты заземленной металлической сеткой или заземленной металлической оболочкой, подсоединенными к системе уравнивания потенциалов. В качестве дополнительной защиты для нагревательных элементов рекомендуется использовать УЗО на ток до 30 мА.
Не допускается использовать для саун, ванных и душевых помещений системы местного уравнивания потенциалов [4].
3 В зданиях, где водоснабжение ванных, душевых и сантехкабин осуществляется ответвлениями в неармированных пластмассовых трубах от распределительной сети, проводящие элементы водопроводной системы: краны, смесители, полотенцесушители, вентили и другие детали, выполненные из металла, не рассматриваются как сторонние проводящие части и не подлежат включению в систему дополнительного уравнивания потенциалов [6].
7 В сантехкабинах для обеспечения электробезопасности защитные контакты розеток, установленных снаружи на сантехкабинах, следует подключать к системе дополнительного уравнивания потенциалов, а светильник в туалете раздельного санузла должен быть класса защиты II, как и в зоне 2 ванной комнаты [6].
Проблема в том, что в с/у нет электрооборудования, за исключением светильника класса защиты II. Иногда устанавливают рукосушители. Но, даже если у нас будет рукосушитель, как обосновать ДСУП в с/у? Неужели он так опасен?
Смысл ДСУП заключается в том, чтобы соединить доступные прикосновению открытые проводящие части электрического оборудования и сторонние проводящие части строительных конструкций и систем заземления для выравнивания потенциала. Для выравнивания потенциала должно быть как минимум 2 разные части.
Можно попытаться обосновать ДСУП в с/у отсутствием основной системы уравнивания потенциалов в здании, а в старых жилых домах вряд ли она выполнена. Однако, где в нормативных документах имеется данное требование? Придет время, при реконструкции дома сделают все как положено.
Некоторые придерживаются мнения, что лучше перестраховаться, однако, с учетом того, что госстройэкспертиза любит все порезать, поэтому приходится проектировать все четко в соответствии с нормативными документами. Я недавно рассказывал, как мне дали замечание, что я отключил кондиционер при пожаре…
«Новости Электротехники»Вопрос: Требуется ли устройство дополнительной системы уравнивания потенциалов в санузле, где только унитаз и умывальник? В этом помещении проходят стояки водопровода и канализации, из электрооборудования на стене под потолком установлен светильник класса защиты 2.
Людмила Казанцева, УИЦ НИИПроектэлектромонтаж АНО
Выполнение дополнительной системы уравнивания потенциалов в санузле в указанных условиях не требуется.
А при каких условиях нужно делать дополнительную систему уравнивания потенциалов в санузле?
Основные ТНПА по проектированию ДСУП:1 ТКП 45-4.04-149-2009 (Системы электроснабжения жилых и общественных зданий) (РБ).
2 ТКП 339-2011 (Вместо ПУЭ) (РБ).
3 ГОСТ 30331.11-2001 (Требования к специальным электроустановкам. Ванные и душевые помещения) (РБ).
4 ПУЭ.
5 СП 256.1325800.2016 (Электроустановки жилых и общественных зданий. Правила проектирования и монтажа) (РФ).
6 ТЦ №23-2009 (Об обеспечении электробезопасности и выполнении системы дополнительного уравнивания потенциалов в ванных комнатах, душевых и сантехкабинах) (РФ).
7 ГОСТ Р 50571.7.701-2013 (Помещения для ванных и душевых комнат) (РФ).
Про ДСУП и ТЦ №23-2009:
Продолжение следует… в любом случае будет интересно.
Советую почитать:
Вы можете пролистать до конца и оставить комментарий. Уведомления сейчас отключены.
Коробка уравнивания потенциалов — подробно и доходчиво
В наших квартирах и домах, производственных помещениях и офисах, где мы работаем, полным-полно металлических корпусов и конструкций, во время одновременного прикосновения к которым человек может попасть в зону разности потенциалов. Чтобы такого не произошло потенциалы надо уравнять. Как это сделать практически? Соединить все имеющиеся в здании токопроводящие элементы. Такая система уравнивания потенциалов (СУП) создаёт безопасную для человека среду. Одним из элементов СУП является коробка уравнивания потенциалов (КУП).
Об этих СУП и КУП поговорим более подробно, но сначала рассмотрим на практических примерах, что представляет собой разность потенциалов в обычных квартирах и откуда она появляется.
Причины
Все мы учили физику и помним, что потенциал сам по себе опасности абсолютно никакой не представляет. Опасаться надо разности потенциалов.
В квартирах разность потенциалов у труб и бытовых электроприборов может возникнуть вследствие следующих обстоятельств:
- Повредилась изоляция провода, и происходит утечка тока.
- В системе заземления возникли блуждающие токи.
- Схема подключения электрического оборудования выполнена неправильно.
- Проявляется статическое электричество.
- Электрические приборы неисправны.
Опасность
Помните со школы? Любой металлический предмет проводит электрический ток. В наших домах подобные предметы повсюду. Это – трубы центральной отопительной системы, холодного и горячего водопровода; батареи и полотенцесушитель; короб вентиляции и водосток; металлический корпус любого электроприбора.
В общедомовых коммуникациях металлические трубы между собой взаимосвязаны. Рассмотрим простой пример. У нас есть ванная комната, в которой рядом расположены батарея отопления и душевая кабинка. Если вдруг между этими двумя элементами возникает разность потенциалов, а человек в одно время прикоснётся и к батарее, и к душевой кабинке, будет крайне опасно в плане поражения током. В данном случае тело человека сыграет роль перемычки, по которой потечёт электрический ток. Путь его протекания нам известен из законов физики – от потенциала с большим значением к меньшему.
Ещё один типичный пример, если разные потенциалы возникают на трубах водопровода и канализации. Когда на водопроводной трубе появляется токовая утечка, есть вероятность поражения человека во время купания в ванной. Это произойдёт в том случае, если человек стоит в ванной с водой, при этом открывает слив и касается рукой водопроводного крана. Чтобы подобных проблем не возникало, необходимо уравнивание потенциалов.
Ситуация, когда на трубах в жилом доме присутствует напряжение, показана в этом видео:
Виды
Для того чтобы уравнивать потенциалы существует две системы, о каждой из них мы поговорим более подробно.
Уравнивание основное
Главной считается основная система уравнивания потенциалов, в сокращённом виде она называется ОСУП. По сути, эта система представляет собою контур, объединяющий несколько элементов:
- наиболее важный – главную заземляющую шину (ГЗШ), именно на ней соединяются все остальные элементы;
- всю металлическую арматуру многоэтажного жилого дома;
- молниезащиту здания;
- отопительную систему;
- детали и элементы лифтового хозяйства;
- короба вентиляции;
- металлические трубы водоснабжения и отвода воды.
Каждое здание имеет вводное распределительное устройство (ВРУ), в нём устанавливают главную заземляющую шину (ГЗШ). Она подключается на контур заземления при помощи стальной полосы.
Раньше не нужно было беспокоиться, все металлические элементы объединялись, и не возникало предпосылок для разных потенциалов. Если и появлялся какой-то потенциал на трубе, по пути наименьшего сопротивления он спокойно уходил в землю (мы ведь помним, что металл – это отличный токопроводник).
Сейчас ситуация изменилась, многие жильцы во время ремонтных работ в квартирах меняют металлические водопроводные трубы на полипропиленовые либо пластиковые. За счёт этого общая цепочка разрывается, батареи и полотенцесушители остаются без защиты, потому что пластик не обладает проводящей способностью и не связан с заземляющей шиной. Представьте, что у вас остались металлические трубы, а сосед снизу всё поменял на пластик. При появлении потенциала на ваших трубах ему некуда уходить, путь в землю прерван пластиковыми трубами соседа. Таким образом и происходит возникновение разности потенциалов.
Есть у основной системы небольшая проблема. В многоэтажных зданиях коммуникационные пути очень протяжённые, за счёт этого увеличивается сопротивление проводящего элемента. В величине потенциала на трубах первого и последнего этажей будет ощутимая разница, а это уже представляет собой опасность. Поэтому создаётся дополнительная система уравнивания потенциалов, она монтируется на каждую квартиру индивидуально.
Дополнительное уравнивание
Дополнительная система уравнивания потенциалов (сокращённое название ДСУП), монтируется в санузлах, в ней объединяются такие элементы:
- металлический корпус душевой кабинки или ванная;
- вентиляционная система, когда её выход в ванную выполнен коробом металлическим;
- полотенцесушитель;
- канализация;
- металлические трубы водопровода, отопления и газового хозяйства.
А вот тут уже понадобится коробка уравнивания потенциалов. К каждому из вышеперечисленных объектов подсоединяется отдельный провод (одножильный, материал исполнения – медь), его второй конец выводят и подсоединяют в КУП.
Выполнение монтажа
КУП различается в зависимости от того, как конструктивно выполнено здание и куда будет монтироваться сама коробка:
- в сплошную стену;
- в полую стену;
- на стенную поверхность (открытый способ установки).
Представляет собой корпус, выполненный из пластика, внутри которого располагается главный элемент – заземляющая шина. Она изготавливается из меди и имеет сечение не менее 10 мм2.
К этой шине через имеющиеся на ней разъемы подсоединяются медные провода от объектов водопроводной, отопительной и газовой систем; от находящихся в помещении электроприборов, а также от розеток и осветительных приборов, установленных в ванной комнате.
Подключение проводов к перечисленным элементам происходит за счёт болтовых соединений либо хомутов. Иногда используют специальные контактные лепестки, в этом случае металлическая связь между защищаемым элементом и проводом буде особенно прочной. Чтобы система уравнивания потенциалов в опасных ситуациях работала, нужен надёжный контакт. Поэтому место на трубах, где будет устанавливаться хомут, нужно зачищать до металлического блеска.
Внутренняя шина отдельным медным проводом, называемым защитным РЕ-проводником, соединяется с вводным квартирным щитком, а уже через него подключается непосредственно к ГЗШ. Сечение РЕ-проводника должно быть не менее 6 мм2. Важное условие, если вы решите проложить этот провод в полу, он не должен пересекаться с другими кабелями.
Такая коробка является как бы промежуточным звеном между всеми заземляющимися элементами и вводным щитком. Очень удобно, что от каждого элемента достаточно протянуть проводок только на КУП, а не к общему квартирному щиту.
Когда разводка выполнена пластиковыми трубами, в КУП подсоединяются провода от водопроводных кранов и смесителей.
Перед тем, как монтировать СУП, необходимо узнать, как в доме выполнено заземление. Если по системе TN-C (когда в один провод совмещаются защитный проводник РЕ и рабочий ноль N), выполнять уравнивание нельзя. Это вызовет опасность для других соседей, если у них такой системы нет.
Требования
При монтаже КУПа необходимо придерживаться некоторых требований и правил:
- Её монтаж в ванных комнатах и санузлах обязателен. Во-первых, в этих помещениях расположено много металлических корпусов и поверхностей. Во-вторых, здесь имеется немалое количество электрических приборов. В-третьих, в этих комнатах всегда высокая влажность.
- Устанавливается коробка в том месте, где проходят сантехнические стояки.
- Обязательно подключение всего электрического оборудования, к которому имеется открытый доступ (это, прежде всего, корпуса водонагревательных бойлеров, стиральных машин), а также сторонних проводящих элементов.
- Доступ к КУП должен быть свободным.
- Установка КУП запрещена, когда в доме заземление смонтировано без заземляющего проводника (методом зануления).
- ДСУП запрещается подключать шлейфом.
- ДСУП по всей длине, начиная от КУП в санузле и до самого вводного щитка, нельзя разрывать. Запрещается монтировать в этой цепи любые коммутационные аппараты.
Напоследок хотелось бы сказать, не путайте понятия уравнивание и выравнивание разных потенциалов. Уравнять – значит соединить проводящие элементы электрически, чтобы сделать их потенциалы равными. А выровнять – это снизить разность потенциалов на полу или поверхности земли (шаговое напряжение).
Если в электричестве у вас опыта маловато, то не беритесь сами за такую работу, доверьте её профессионалам. Кроме всего прочего, специалист по окончании монтажных работ должен ещё померить сопротивление заземления, и проверить наличие цепи между заземляющими элементами.
Уравнивания потенциалов. Виды и применение. Установка
Когда в ванной бьет током от металлических труб, то решить такую проблему можно установив специальную защиту для металлических объектов, которая называется система уравнивания потенциалов.
Обычно в новостройках планируют и выполняют такие защитные системы от поражения током. Но в старых домах такой вариант не всегда работает. Разберемся, что представляет собой система уравнивания потенциалов (ее сокращенная аббревиатура СУП), ее виды, и как можно самому ее сделать.
Назначение
Выясним, необходима ли система уравнивания потенциалов в обычной квартире. Все предметы, выполненные из металла, проводят электрический ток. Это нам известно из школьных уроков по физике. В наших квартирах опасными местами являются отопительные трубы, а также трубы водоснабжения, водосточная труба, водопровод, полотенцесушитель в ванной, коробы вентиляции.
Все металлические коммуникации в доме связаны друг с другом. При возникновении разности потенциалов между некоторыми объектами из металла, например, ванной и радиатором отопления, касание человека сразу этих двух объектов может привести к удару электрическим током. Это происходит потому, что тело выступает в качестве перемычки между батареей и ванной, поэтому ток протекает по телу человека от объекта, имеющего больший потенциал, к объекту с наименьшим значением потенциала.
Подобный случай опасности – это появление разности потенциалов на трубах канализации и водопровода. При возникновении утечки тока на водопроводных трубах, когда человек моется в ванной, будет высока вероятность удара током при касании включенного крана. Вода проводит ток от водопровода к канализации, а вы замыкаете своим телом эту цепь.
Чтобы исключить наличие такой опасности, необходимо уравнивание потенциалов с помощью специальной системы, установленной в квартире.
ВидыСуществует два вида систем уравнивания потенциалов:
- Основная (ОСУП).
- Дополнительная (ДСУП).
Это главная система уравнивания потенциалов, представляющая собой контур, объединяющий следующие элементы этой системы:
- Заземлитель.
- ГЗШ – главная заземляющая шина. Она расположена на вводе в здание.
- Металлические части арматуры жилого дома.
- Короба вентиляционной системы.
- Трубы водопровода из металла (горячее и холодное водоснабжение).
- Защита от молнии.
В ранние времена при объединении всех этих частей не было опасности появления разности потенциалов. Но сегодня положение в корне изменилось, так как хозяева многих квартир заменяют прогнившие металлические трубы пластиковыми, либо полипропиленовыми, которые не проводят электрический ток. Пластиковые трубы разрывают цепь, в результате появляется разность потенциалов между разными металлическими деталями в ванной.
У основного вида системы имеется существенная проблема, которая заключается в том, что на значительной протяженности труб, например, в 12-этажном доме, электрический потенциал одной и той же трубы на первом и последнем этаже будет иметь большое отличие. Это приводит к опасной ситуации. Поэтому необходима вспомогательная система, о которой расскажем ниже.
ДСУПЭта система является дополнительной, и располагается в ванной комнате. Она включает в себя такие элементы:
- Корпус душевой кабины, либо ванны.
- Сушка для полотенца.
- Трубы: газовые, водоснабжения, отопления.
- Канализационная система.
- Короб вентиляционной системы.
Каждый элемент этой системы соединяется отдельным проводом с медной жилой. Второй конец этого провода выводят в специальную коробку (КУП).
Существуют определенные требования к созданию ДСУП по правилам ПУЭ:
- Нельзя подключать составные части ДСУП шлейфом.
- Запрещается выполнять ДСУП, при условии, если в квартире не установлен контур заземления.
- Дополнительная система не должна разрываться на своем протяжении от коробки КУП до квартирного щита. В цепи нельзя устанавливать аппараты коммутации.
Если у вас нет такого защитного контура, как уравнивание потенциалов, расскажем ниже, как его можно выполнить своими силами.
Установка системы уравнивания потенциаловУстановить вспомогательную систему по выравниванию потенциалов не составляет большой сложности. Ее называют местной системой. Но такую работу лучше выполнять при проведении ремонта в квартире, так как необходимо проводит провод до щита от коробки КУП под полом, а это связано с нарушением покрытия пола, и сопутствующих ремонтных работ.
Для начала монтажа готовят некоторые материалы по следующему перечню:
- Клеммная коробка в комплекте с шиной из меди (ШДУП).
- Медные провода, состоящие из одной жилы. Площадь сечения проводов должна быть от 2,5 до 6 мм2, марки ПВ-1.
- Крепежные элементы: болты, хомуты, фиксирующие лепестки. Они необходимы для соединения проводов всей системы уравнивания с трубами и металлическими частями.
С таким комплектом элементов можно начинать установку ДСУП. Сначала составляют схему соединений, чтобы выполнить правильное уравнивание потенциалов. На схеме также изображают места прохода провода от коробки КУП до шины заземления в квартирном щите. На рисунке показан один из примеров проекта.
Далее, готовятся к подключению сами коммуникации, то есть, зачищаются место контакта хомута с трубой, до появления металлического блеска. Это необходимо для надежности соединения. В опасной ситуации уравнивание потенциалов сработает как положено.
Затем подключают провода к каждому элементу системы. Если вы уверены в том, что не произойдет повреждения провода, то достаточно сечения провода размером 2,5 мм2. Но если имеются какие-либо сомнения по этому поводу, то лучше применить провод на 4 мм2. Все проводники проводят в коробку и выполняют надежное соединение с шиной.
Клеммная коробка для ванной комнаты должна иметь степень защиты не менее IР54. от шины коробки должен быть выведен провод сечением 6 мм2 до квартирного щита. Здесь имеется свое требование в том, что этот провод не должен иметь пересечения с другими кабелями разных линий.
В конце работы провод соединяют с заземляющей шиной щита. На этом монтаж можно считать законченным. Для самоуспокоения можно вызвать квалифицированного электромонтера для проверки работы системы с помощью приборов, а также визуальным осмотром.
Ограничения монтажа СУПУстановку СУП рекомендуется производить во время строительства здания. Но есть некоторые ограничения по ее использованию в уже построенных домах, в которых заземление выполнено по системе ТN-С, с объединенным РЕN проводником. В таких домах запрещается выполнять уравнивание потенциалов. Иначе, во время обрыва нулевого провода создается опасность удара электрическим током жильцов других квартир, в которых нет ДСУП. Чаще всего такое ограничение распространяется для многоэтажных домов старого фонда.
Эту проблему разрешают путем перехода на заземление по системе ТN-С-S. Для этого в распределительном щите дома на главной шине заземления проводник РЕN разъединяют на РЕ и N проводы, осуществляют подключение заземляющего контура и подключают его к основной шине заземления медным проводником.
В настоящее время имеется тенденция замены металлических труб на пластиковые, которые не требуют их подключение к СУП. Если у вас уже имеется дополнительное уравнивание потенциалов металлических труб, а вы решили заменить трубы на пластиковые, то это приведет к разрыву электрической связи с шиной заземления остальных элементов, изготовленных из металла. Это сделает их опасными для человека при прикосновении одновременно к нескольким частям.
Новые правила и нормы строительства направлены на соблюдение правильности установки уравнивания потенциалов. Эту систему подвергают осмотру, проверяют по проекту перед сдачей дома. Электрическая безопасность создается при выполнении электрических соединений всех металлических частей, доступных для касания человека, с основной заземляющей шиной путем РЕ проводов.
Основная система дополняется местными системами уравнивания в местах с большой опасностью удара электрическим током. Нельзя забывать, что при установке СУП должна быть обеспечена надежная связь между элементами системы, которые подключены по радиальной схеме. При этом сечение провода должно быть не менее рекомендованного значения.
Уравнивания потенциалов молниезащитной системыПри ударе молнии возникает большая сила тока и скорость его нарастания. Из-за этого появляется разница потенциалов больше, чем от утечки тока в сети. Поэтому для создания защиты от молнии необходимо выровнять потенциалы.
Чтобы при ударе молнии не было неконтролируемых замыканий, нужно непосредственно соединить электрические устройства, металлические элементы, заземление, защитную систему от молнии с устройствами защиты. Проводники всей системы соединяются с уравнивающей шиной, которая должна быть доступна для целей испытания, она соединяется с заземляющим контуром. Большие здания обычно имеют несколько таких шин. При этом все они соединены друг с другом.
Система уравнивания потенциалов молниезащиты осуществляется на вводе в здание, и в местах, где нельзя соблюсти безопасные расстояния, например, на уровне земли, либо в подвале.
В бетонном здании, либо с каркасом из металла или имеющем молниезащиту отдельного исполнения, уравнивание молниезащиты выполняется только на уровне грунта. В высоких зданиях выше 30 метров, на каждые 20 метров делается уравнивание потенциалов молниезащиты.
Молниепроводящие детали располагают на безопасном расстоянии от СУП, во избежание импульсных перекрытий. Если такое расстояние нельзя обеспечить, то создаются вспомогательные связи между молниеотводом, молниеприемником и СУП. При этом учитывают фактор того, что вспомогательные связи дают возможность захода высокого потенциала в здание.
Похожие темы:
Система уравнивания потенциалов | Электрик
В нашем доме находятся различные металлические установки и предметы быта, кухонные мойки, металлические ванны, полотенцесушытели и батареи отопления, а также многое другое.
Все эти предметы, по законам физики, способны проводить электрический ток. Грубо говоря, их можно назвать проводниками.
В обычном состояние все ети проводники, как и любые другие проводники имеют равномерное распределение электронов, как положительных, так и отрицательных, по всей своей внутренней структуре.
Если подключить проводник к оборудованию, которое создает на одном своем полюсе недостаток электронов, а на другом своем полюсе их избыток, то все электроны нашего проводника начнут направленное движение, чтобы выровнять этот недостаток и избыток.
То есть вернутся опять в «обычный» режим. Такое направленное движение электронов и называется электрическим током, а создаваемый на полюсе проводника избыток или недостаток электронов называется отрицательным и положительным электрическим потенциалом.
По законам физики, каждый проводник обладает каким то электрическим потенциалом.
Например, если между потенциалом батареи отопления и корпусом стиральной машыны есть разница то такую разницу можно считать напряжением.
И хоть эти вещи не находятся фактически под фазой, все же в действительности, по множеству причин, разница потенциалов может иметь опасно высокое напряжение.
К таким причинам можно отнести, например, повреждение изоляции, статическое электричество и блуждающие и циркулирующие токи систем заземления.
Чтобы решать эту проблему и безопасно пользоваться бытовой техникой и ванной, применяют систему уравнения потенциалов, ее суть довольно проста, если токоведущие части имеют непосредственное электрическое соединение, то их потенциал всегда одинаков, и напряжение между ними не возникнет ни при каких обстоятельствах.
Поэтому к системе уравнения потенциалов подключают все металлические предметы, трубы, щитки, короба и бытовую технику с металлическим корпусом. Все ети предметы подключаются к главной заземляющей шине.
Система уравнения потенциалов бывает:
- основная система уравнения потенциалов — ОСУП
- дополнительная система уравнения потенциалов — ДСУП
ОСУП включает в себя: контур заземления, главную заземляющую шину, сетки защитных проводников (РЕ) и сами проводники уравнения потенциалов.
Следует помнить что соединять защитные проводники (РЕ) с проводниками N — запрещается!
Схема подключения к заземляемым элементам, конструкциям и инженерным сетям здания должна быть радиальной, то есть на каждую заземляемую часть строения должен быть свой проводник уравнивания потенциалов. Подключать шлейфом РЕ-проводники строго запрещается!
А самое главное требование — не должно быть никаких коммутационных элементов, должна быть обеспечена полностью непрерывна защита проводников.
ДСУП — дополнительная система уравнения потенциалов нужна для того чтоб обеспечить дополнительную электробезопасность в помещениях с повышенной опасностью, в ванной комнате или душевой.
ДСУП состоит из монтажной коробки уравнения потенциалов, внутри которой находится латунная шина и самих соединительных проводников уравнения потенциалов, как правило это медные провода сечением 2.5 — 6мм.
К ДСУП подключают отопление, водопровод, ванную, душевую, а также все розетки в ванной и других влажных помещениях.
Так как на проводники действуют законы сопротивления — проводников большой протяжности быть не должно. Другими словами, электрический потенциал железной трубы на вводе в помещение и на девятом этаже имеет возможность очень отличатся и главная система уравнивания потенциалов становится все наименее действенной по мере удаления от ГЗШ.
Потому в любой жилплощади здания создается отдельная, вспомогательная система выравнивания потенциалов. Ее проводники подключаются к шине РЕ в квартирном щитке.
Система уравнивания потенциалов — это чрезвычайно важная и нужная вещь, она обладает сопротивлением, хотя и не огромным.
Поэтому, когда по одной ее части проходит электрический ток, к примеру, при срабатывании защитного прибора либо пробое, то и другая часть заземляющего проводника, та через которую ток даже не проходил также окажется под напряжением. Данное напряжение имеет возможность вызвать возникновение циркулирующих токов, действие которых фактически не прогнозируемо. Чтоб этого не произошло, объединяют все подлежащие заземлению металлические корпуса устройств и легкодоступные для прикосновения системы здания, также железные трубопроводы, ванны и душевые.
Когда заземление окажется под напряжением, под ним станут и все элементы, которые доступны для прикосновения, что автоматически понизит возможность поражения электрическим током.
Из этого всего возможно прийти к выводу, что система выравнивания потенциалов считается довольно важным методом защиты при косвенном прикосновении и для обеспечения электробезопасности ее непременно необходимо организовывать при ремонте и модернизации квартирной проводки.
Системы уравнивания потенциалов — основная (ОСУП) и дополнительная (ДСУП). Заземление :: BusinessMan.ru
Со временем здания приобретают все более широкую и усложненную систему электрооборудования. Тем самым потребители с низким вольтажом могут получить больший урон от перенапряжений, вызванных грозой и возникающих из-за воздействия электрических импульсов и уменьшения разделяющего опасного пространства между электрическими предметами и молниеприемником. Объемная система электропроводящих сетей организуется информационным снабжением, антенными конструкциями, коммуникациями централизованного отопления, водоподводящими, газовыми и силовыми системами. Единственная молниезащита при воздействии электромагнитного импульса не способна предотвратить повреждение достаточно слабого оснащения. Поэтому должна формироваться сеть общей молниезащиты, и в первую очередь основная система уравнивания потенциалов.
Для чего применяется
Уравнивание потенциалов используется для обеспечения выравнивания во всех металлических частях здания, связанных между собой, то есть для формирования эквипотенциальной поверхности. В этом случае при попадании в дом повышенного потенциала на всех конструкциях из металла он увеличивается синхронно, за счет чего не развивается опасная разница в напряжении и не образуется искрение и прохождение опасных токов.
Соединяющие элементы
Важным защитным мероприятием является создание главной системы уравнивания потенциалов. Соединяется ею заземляющая основная шина, основная магистраль заземления, защитная основная магистраль и проводящие элементы, к которым относятся:
- арматурные детали конструкций с железобетонным основанием;
- элементы зданий из металла, климатические системы, централизованное отопление;
- стальные трубопроводы системного питания.
Чаще всего у системы выравнивания потенциалов присутствует только один способ вывода. В помещении распределительного элемента монтируется главная шина на максимально близком расстоянии от точки введения.
Система молниезащиты
Из-за быстроты нарастания тока и его большой силы при ударе молнии создается огромная разница потенциалов, намного превышающая ту, которая возникает в связи с утечкой тока. Поэтому требуется выравнивание потенциалов для защиты от влияния токов молний.
Для предотвращения неконтролируемого замыкания должны быть сторонне или напрямую совмещены молниезащитная конструкция, заземляющая система, оснастка из металла, электроустановки с защитными механизмами.
Шина уравнивания потенциалов с открытым доступом для проверочных работ должна иметь соединение с уравнивающей системой. Также шина обладает соединением с заземлением. В больших зданиях их может быть несколько, если они будут обладать соединением между собой.
Выравнивание потенциалов в молниезащитной системе осуществляется в месте попадания проводников в помещение и там, где нарушены безопасные дистанции, на уровне почвы или в подвале.
Дом, построенный с использованием стального каркаса или железобетонного основания либо с отдельным помещением для внешней молниезащиты, должен обладать уравниванием потенциалов на уровне грунта. В домах с высотой более 30 м оно выполняется каждые 20 м.
Молниепроводящие детали устанавливаются на безопасном расстоянии для предотвращения появления импульсных реакций. При невозможности соблюдения безопасной дистанции системы уравнивания потенциалов, устройство отведения молнии и приемник формируют между собой дополняющие связи. Стоит отметить, что они способны привести к занесению в строение повышенного потенциала.
Дополняющее устройство
Создается дополнительная система уравнивания потенциалов, ПУЭ которой определяет форму и применение, в точках расположения электрооборудования, в которых имеющиеся условия могут быть опасными, и в случае, если нормы свидетельствуют о необходимости в ней. Она образует связь между всеми частями имеющегося оборудования и сторонними проводниками, которые находятся рядом с ними.
Типичными помещениями и объектами, в которых должны использоваться дополняющие меры безопасности, являются антенное оборудование, объекты молниезащиты, сооружения удаленной связи, участки с повышенной взрывоопасностью, госпитали, фонтаны, аквапарки, ванные комнаты. Компания, которая занимается выполнением монтажных работ, должна осуществлять их в соответствии с указаниями ПУЭ-7.
Потенциалы защиты от молний и оснастка
Должно производиться соединение системы защиты от молний и деталей оснастки, к которой причисляются воздуховоды климатических и вентиляционных устройств, крановые каркасы, направляющие элементы лифта, трубопроводы таких систем, как пожаротушение, теплообеспечение, газо- и водоподведение. При наличии возможности каждая металлическая конструкция соединяется с шинами уравнивания. Электропроводящие трубы могут выступать в роли соединительных линий (исключение составляет газопровод).
Если имеется изолированный участок на водо- и газопроводе, используются для шунтирования проводники системы уравнивания потенциалов. Специальное соединение с устройством защиты от молний не нужно для подземных трубопроводов из металла, находящихся рядом с заземлением. То же самое касается железнодорожных рельсов. Если без объединения не обойтись, то предварительно оно согласовывается с эксплуатирующей компанией.
Заземление
Работает заземляющее повторное устройство при помощи двух вертикальных электродов с длиной не менее 5 м, между собой они скреплены горизонтальным заземлителем. В роли последнего выступает стальная полоса, также она применяется для формирования проводника, соединяющего ГЗШ и дополнительный заземлитель. Полоса должна быть не меньше 4 мм толщиной с площадью поперечного сечения 75 мм2. Нормирование сопротивления повторного заземлителя отсутствует.
Сечение питающего кабеля оказывает влияние на подбор проводника выравнивания потенциала, он не должен быть меньше половины сечения кабеля. Наибольшее распространение приобрела проводка ПВ1 и стальная полоса, также используется одножильный кабель. Специальные сжимы зачастую применяются при ответвлении магистрали при помощи провода.
Техническое оборудование и молниезащита
В соответствии с тезисами ПУЭ-7 и при соблюдении границ сечения проводников выполняются все соединения для выравнивания потенциалов конструкций защиты от молний. Должны разделяться соединения непосредственные и осуществляемые через искровые разделяющие промежутки.
Система молниезащиты может иметь непосредственное объединение со следующими устройствами:
- заземляющие элементы системы защиты от высокого напряжения сооружений охранного типа;
- антенные приспособления;
- линии заземления, находящиеся под землей в удалении от систем коммуникации и защиты от превышения напряжения;
- заземление силовых конструкций, мощность которых превышает 1 кВт, при этом должна отсутствовать возможность занесения в заземлители высокого потенциала;
- предохраняющие связи в сетях типа ТТ для защиты от удара током при косвенных контактах.
При проведении в металлических трубах или экранизации информационных либо силовых линий дополнительная система уравнивания потенциалов не нужна.
Искровые промежутки
Контрольные испытания должны осуществляться при получении доступа к искровым разъединительным пространствам. Благодаря правильной проектировке и установке механизма внутренней защиты от молний минимизируются повреждения, вызванные разностью потенциалов и импульсами перенапряжения.
Соединение через искровые промежуточные разделения осуществляется для следующих элементов:
- заземление измеряющих систем при условии отдельного проектирования;
- установки, защищенные от утечки тока и имеющие антикоррозионную катодную защиту;
- обратный провод тягового элемента постоянного тока, а также переменного при отсутствии возможности выполнения непосредственного объединения по сигнально-техническим доводам;
- вспомогательные заземляющие детали защитного отключения, которое срабатывает при опасном напряжении.
Установка
Во время строительства здания должен выполняться монтаж СУП, так как есть некоторые сложности при использовании в готовых строениях. Дополнительная коробка уравнивания потенциалов запрещена к применению в зданиях, имеющих заземление вида TN-C. При несоблюдении данного правила во время разрыва нулевого провода есть вероятность поражения током жильцов, которые не устанавливали ДСУП. Относится данное ограничение в основном к старому многоэтажному жилому фонду.
Заземляющая система другого типа позволяет избавиться от такой проблемы: для этого выполняется заземляющий контур и присоединяется медной проводкой к защемляющей главной шине.
Пластиковые трубы
Сегодня имеет достаточное распространение проведение коммуникаций с использованием пластмассовых труб, для которого не нужно объединение с системой уравнивания. При этом, если в существующей ДСУП заменить трубы из металла на пластиковые, отличающиеся токопроводящими свойствами, возникнет нарушение связи между металлическими частями в помещении (полотенцесушитель, батареи) и заземляющей шиной, из-за чего они становятся опасными при одновременном касании.
При создании коммуникаций с помощью труб из пластика объединение с системой уравнивания осуществляется с применением металлических гребенок, кранов и обратных клапанов для закрепления проводников. При наличии диэлектрических вставок в металлических трубах они добавляются к главной системе после вставок внутри строения.
Что нужно знать
В соответствии со строительными правилами и нормами, сегодня уделяется повышенное внимание грамотной установке системы уравнивания потенциалов. В первую очередь осуществляются при сдаче здания в эксплуатацию осмотр и проверка на соответствие проекту. Создание электрического объединения всех проводящих элементов, доступных для касания, с помощью специальных проводников обеспечивает должную электробезопасность. В качестве дополнения выступает коробка уравнивания потенциалов в местах с высокой возможностью поражения током.
Стоит учитывать то, что ДСУП может создаваться только в зданиях, которые имеют систему заземления с раздельным прокладыванием проводников N- и PE-типа.
Между частями СУП должна устанавливаться металлическая прочная связь, если они подключены в соответствии с радиальной схемой и необходимым сечением защитного проводника.
Дополнительная система уравнивания потенциалов в ванных и душевых помещениях
Дополнительная система уравнивания потенциалов в ванных и душевых помещениях
Рубрика: Статьи ‡Довольно часто при выполнении электромонтажных работ в ванной или душевой комнате возникают вопросы о дальнейшей безопастной эксплуатации этих помещений. При разработке проектов электроснабжения квартиры, электроснабжения дома, электроснабжения административного здания учитываются все требования номативных документов, но есть некоторые моменты, на которые нормативные документы четкого ответа не дают. Поэтому в этой статье мы постараемся ответить на вопросы касательно электробезопастности ванных и душевых комнат.
Вопрос №1. Необходимо ли присоединять к шине дополнительной стистемы ураснивания потенциалов (ДСУП) металлических смесителей холодной и горячей воды, отопительный металлический радиатор, если трубы присоединяемые к ним пластмассовые?
Ответ: действительно, на сегодняшний день все большее количество людей при ремонтах в ванных меняют старые металлические трубы, которые являются хорошими проводниками тока, на новые пластмассовые, которые являются диэлектриком. Поэтому если в ванной комнате краны холодной и горячей воды и отопительный металлический радиатор установлены на пластмассовых трубах, то, пренебрегая электропроводностью воды, можно считать, что они изолированы и присоединять их к шине уравнивания потенциалов не требуется.
Вопрос №2. Необходимо ли выполнять уравнивание электрических потенциалов и заземление металлических корпусов ванн в жилых домах в случае, если водопроводные сети выполнены пластмассовыми трубами.
Ответ: Для ванных и душевых помещений является обязательным требование выполнять дополнительную систему уравнивания потенциалов.
К дополнительной системе уравнивания потенциалов должны быть присоединены все доступные одновременному прикосновению открытые проводящие части стационарных электроприемников, сторонние проводящие части (металлические ванны и душевые поддоны, металлические части строительных конструкций и др.), а также нулевые защитные проводники указанных электроприемников и штепсельных розеток. К дополнительной системе уравнивания потенциалов следует также подсоединить заземленную металлическую сетку или заземленную оболочку замоноличенного в пол нагревательного кабеля, если таковой имеется.
Металлическая ванна или душевой поддон, даже если водопроводные и канализационные трубы выполнены из пластмассы, не являются изолированными поскольку, во-первых, водопроводная вода не является диэлектриком, во-вторых, ванна или душевой поддон могут иметь гальваническую связь с заземленными металлическими частями строительных конструкций.
Поэтому, если отсутствуют стационарное электрооборудование с подключенными к дополнительной системе уравнивания потенциалов зажимами нулевых защитных проводников, к которым можно присоединиться, или другие стационарные металлические предметы, подключенные к дополнительной системе уравнивания потенциалов, к которым можно надежно присоединиться, металлические корпуса ванн и душевых поддонов следует присоединить к РЕ шине (зажиму) квартирного щитка с помощью проводника.
Вопрос №3. Выполняя дополнительную систему уравнивания потенциалов в ванной комнате можно ли эту шину присоединить к РЕ проводнику розетки, устанавливаемой для стиральной машины и не прокладывать провод к РЕ шине на вводе в квартиру.
Ответ: при наличии в ванной комнате коробки с шиной уравнивания потенциалов и розетки для стиральной машины шину уравнивания потенциалов можно присоединять к РЕ проводнику розетки, если такое решение приемлемо с конструктивной точки зрения.
К записи 2 комментария
Оставить комментарий или два
Пожалуйста, зарегистрируйтесь для комментирования.
Белок-супрессор повреждений тихоходок связывается с нуклеосомами и защищает ДНК от гидроксильных радикалов
Благодарим рецензентов за положительную и восторженную оценку данной работы. Получив отзывы, мы предложили стратегию решения проблем, которые были подняты в основных редакциях, и этот план был одобрен редакторами eLife .
К счастью, в ходе нашего анализа структуры и функции Dsup при пересмотре этой работы мы сделали замечательное открытие, что консервативная область Dsup демонстрирует сходство последовательностей с основным консенсусным связывающим нуклеосому доменом белков HMGN позвоночных ( новый рисунок 7A).Важно отметить, что с помощью мутационного анализа мы обнаружили, что HMGN-подобная последовательность в Dsup функционально важна для связывания нуклеосом (новая фигура 7C) и защиты хроматина от гидроксильных радикалов (новая фигура 7D). Эти результаты обеспечивают следующее: (i) веские доказательства того, что связывание Dsup необходимо для защиты ДНК от гидроксильных радикалов; (ii) идентификация ключевой области Dsup, которая важна для связывания с нуклеосомами; и (iii) удивительную и интригующую связь между белком Dsup тихоходки и белками HMGN, которые были обнаружены только у позвоночных.Эти открытия придают этой работе новое захватывающее измерение.
Важно отметить, что эти выводы касаются основных концептуальных вопросов, поднятых рецензентами. Мы также рассмотрели технические вопросы, такие как количественный анализ данных о сдвиге подвижности геля (новый рисунок 2 — добавление к рисунку 1), переименование Hypsibius dujardini в Hypsibius instanceplaris и пересмотр материалов и методов. .
Существенные изменения:
1) Наиболее критический вопрос для пересмотра касается отсутствия естественной системы отсчета для интерпретации наблюдаемого влияния Dsup на связывание нуклеосом и защиту от расщепления гидроксильным радикалом.Какова стехиометрия Dsup по отношению к нуклеосомам у тихоходок? Если Dsup защищает ДНК тихоходок от повреждений, может показаться, что количество молекул Dsup должно быть в том же порядке, что и количество нуклеосом. Авторы должны прокомментировать относительное содержание Dsup и нуклеосом в клетках. Эта информация важна для интерпретации результатов, что высокие отношения Dsup к нуклеосомам необходимы для защиты от расщепления в экспериментах авторов. Относительная стехиометрия может быть в литературе по тихоходкам, если так, авторам следует процитировать и обсудить.
Что касается этого вопроса, на дополнительном рисунке 10 (см. Также дополнительные таблицы 4 и 5) Hashimoto et al., 2016 показан гель SDS хроматина (Ch) и растворимых ядер (NS) фракций из R. varieornatus . Пептид Dsup находится в полосе B1, которая является основной полосой белка во фракции хроматина, которая не видна в растворимой ядерной фракции. Также можно увидеть, что Dsup колокализуется с хроматином у эмбрионов R. varieornatus с помощью иммунофлуоресценции (дополнительная фигура 12).Однако эти методы не позволяют количественно оценить количество молекул Dsup на нуклеосому у тихоходок. Такие данные пока недоступны, и, учитывая проблемы, связанные с анализом чистой ткани тихоходок, может пройти много лет, прежде чем такие данные будут получены. Тем не менее, Dsup достаточно распространен, чтобы его можно было идентифицировать во фракции хроматина и четко визуализировать с помощью иммунофлуоресценции. Безусловно, это было бы поводом для беспокойства, если бы его нельзя было обнаружить у тихоходок. Поскольку было бы технически чрезвычайно сложно получить качественные количественные данные по тихоходкам, мы сочли, что было бы невозможно получить такие данные для этой статьи или сделать значимый комментарий к существующим данным.К счастью, редакторы eLife согласились с этой оценкой.
2) В то время как рукопись документирует очень интересную и потенциально новую функцию отличительного белка, связывающего нуклеосомы, текущий уровень понимания механизмов ограничен. Как Dsups связывается с нуклеосомами, блокируя расщепление гидроксильных радикалов? Покрывают ли Dsups большие или второстепенные бороздки ДНК или фосфатный остов? Какова роль мотива SAGK в контакте с нуклеосомной ДНК? Достаточна ли кривизна ДНК для предпочтительного связывания с NCP по сравнению с линкерной ДНК, или гистоны также участвуют в связывании? Предоставление дополнительного исследования механизмов по некоторым из этих направлений и включение полученных данных необходимо для обоснования принятия текущей рукописи для eLife.
Четыре точки заключаются в следующем.
i) Гидроксильные радикалы реагируют с атомами водорода на основной цепи дезоксирибозы со следующим предпочтением: 5 ‘H> 4’ H> 3 ‘H ~ 2’ H ~ 1 ‘H [Balasubramanian et al., 1998]. Поскольку атомы водорода на атомах углерода 5 ‘и 4’ обнажены в малой бороздке ДНК, похоже, что Dsup блокирует доступ к малой бороздке ДНК. Этот пункт теперь включен в статью следующим образом:
«Поскольку гидроксильные радикалы реагируют в первую очередь с атомами водорода, которые обнажены в малой бороздке ДНК (Balasubramanian et al., 1998), похоже, что Dsup блокирует доступ к малой бороздке ».
ii) Отсутствует «мотив САГК». Dsup высоко обогащен остатками S, A, G и K (> 60% всех аминокислотных остатков в Dsup R. varieornatus ), но они не существуют в качестве мотива SAGK. Чтобы свести к минимуму вероятность того, что читатели могут непреднамеренно подумать, что существует мотив SAGK, мы изменили (в Обсуждении) «… области, богатые SAGK.…» На «… остатки SAGK.…»
iii) Первые два пункта касаются конкретных вопросов, поднятых в пункте 2, за исключением вопроса кривизны ДНК.В более широком плане можно отметить, что дополнительные данные о возможной роли кривизны ДНК не изменят основных выводов статьи. Я был одним из первых BRE для eLife , и я работал в течение трех лет (а затем некоторое время в качестве старшего редактора). На наших обучающих занятиях нам сказали, что eLife не заинтересован в запросе дополнительных экспериментов, которые не меняют основных выводов статьи. Насколько я понимаю, эта политика не изменилась.
iv) Наконец, мы рассмотрели механизм связывания Dsup с нуклеосомами. Эти эксперименты описаны более подробно ниже в ответе на основные пункты 4 и 5. Вкратце, мы идентифицировали консервативную область Dsup, которая связана с областью связывания нуклеосомы белков HMGN позвоночных. Мутация этой области Dsup приводила к снижению или почти полной потере активности связывания нуклеосом и защиты гидроксильных радикалов. Интересно и интригующе, что Dsup может связываться с нуклеосомами с помощью механизма, который связан со связыванием белков HMGN позвоночных с нуклеосомами.
3) При описании рисунка 2А авторы заявляют: «Эти эксперименты показали, что Rv Dsup связывается с более высоким сродством с нуклеосомами, чем со свободной ДНК». Хотя это, по-видимому, так, вывод был бы более сильным, если бы авторы количественно оценили гели и оценили сродство к Dsup-связывающим нуклеосомам и свободной ДНК.
Мы предоставили количественные данные. Для экспериментов по первичному сдвигу геля (как на рисунках 2A и 2B) мы провели количественную оценку трех независимых экспериментов и предоставили графики (среднее +/– стандартное отклонение процентного соотношения ДНК / нуклеосом в сравнении с молярным отношением Dsup: ДНК / нуклеосома) на новом рисунке. 2 — приложение к рисунку 1.Эти результаты еще раз подтверждают выводы. Мы дополнительно предоставили количественный анализ данных сдвига геля на рисунках 5C и 7C. Мы не пытались оценить абсолютное сродство, так как основной задачей было выяснить, связывается ли Dsup с более высоким сродством с нуклеосомами, чем со свободной ДНК (и абсолютное сродство не повлияет на выводы статьи).
4) Активность защиты ДНК (на рис. 6В) действительно интригующие данные. И RvDsup, и HdDsup продемонстрировали существенную защиту ДНК от гидроксильных радикалов, что действительно здорово.Однако TFIIB не является хорошим контролем для такого рода экспериментов, поскольку он не является связывающим нуклеосомы белком, и любой фактор, связывающий нуклеосомы, может обеспечивать некоторую защиту ДНК от повреждений. Следовательно, другой белок, способный связывать нуклеосомы, следует исследовать в качестве сравнительного аналога Dsups. Кроме того, дайте четкое описание протеинового препарата. для анализа защиты гидроксильных радикалов.
Что касается проблемы защиты гидроксильных радикалов и использования TFIIB в качестве контроля, мы сочли, что было бы лучше всего ответить на эти комментарии путем дальнейшего тестирования, требуется ли связывание Dsup с нуклеосомами для защиты от гидроксильных радикалов.Основываясь на данных в представленной статье, формально было возможно, что Dsup обычно защищает ДНК от гидроксильных радикалов с помощью механизма, который не включает связывание с нуклеосомами. Чтобы ответить на этот важный вопрос, мы создали мутантную версию Dsup (мутация M1), которая значительно уменьшила связывание с нуклеосомами (см. Новые рисунки 7A, 7B и 7C). Важно отметить, что затем мы обнаружили, что мутант M1 Dsup серьезно дефектен по своей способности защищать хроматин от гидроксильных радикалов (рис. 7D). Таким образом, мы смогли показать, что опосредованная Dsup защита от гидроксильных радикалов зависит от связывания Dsup с хроматином.
Что касается очистки белка, мы пересмотрели и уточнили материалы и методы, чтобы четко указать точный метод приготовления Dsup для анализа защиты гидроксильных радикалов.
5) Предыдущая работа показала, что С-конца достаточно для связывания с ДНК и входа в ядро, но недостаточно для защиты генома in vivo . Достаточно ли С-конца для связывания нуклеосомы in vitro , или для защиты нуклеосом требуются дополнительные домены, чем требуется для связывания ДНК? Самоагрегируются ли белки Dsup на массивах нуклеосом?
В Hashimoto et al., 2016 (дополнительный рисунок 14), было показано, что C-концевая область Dsup (N-концевая делеция, лишенная N-концевых остатков 2-207 из 445 аминокислотных остатков) может взаимодействовать с ДНК и локализоваться в ядре. Однако в анализе защиты генома, Hashimoto et al., 2016, тестировали только N-концевую область (то есть C-концевую делецию), а не C-концевую область (дополнительный рисунок 17). Таким образом, не было показано, что С-концевой области «недостаточно для защиты генома in vivo».Следовательно, невозможно продолжить работу по пункту 5, как указано, потому что данных о защите генома с помощью С-концевой области не существует.
Важно отметить, что мы, тем не менее, смогли ответить на вопросы, связанные с проблемами, поднятыми рецензентами в этом пункте. Как показано на новой фигуре 7, мы сделали удивительное наблюдение, что область, которая является консервативной между Rv Dsup и He Dsup, демонстрирует поразительное сходство последовательностей с центральной областью ядра белков HMGN позвоночных (новая фигура 7A).Эта область находится рядом с С-концом белков Dsup. Поэтому мы создали C-концевую делецию Dsup (M1; делеция аминокислотных остатков 360-445), а также мутант Dsup с тройной заменой аминокислот с R на E (обозначенный M2) (новая фигура 7A). Затем мы обнаружили, что мутант M1 почти полностью дефектен для связывания с нуклеосомами и что мутант M2 умеренно дефектен для связывания нуклеосом (новый рисунок 7C). Таким образом, мы идентифицировали консервативную область Dsup, которая важна для связывания с нуклеосомами и защиты от гидроксильных радикалов (новая фигура 7D).Интересно, что эта область связана с основным связывающим нуклеосомы доменом белков HMGN позвоночных. Поскольку нуклеосомный связывающий домен HMGN не был обнаружен за пределами позвоночных, действительно довольно неожиданно и интересно найти родственную и функционально важную последовательность у тихоходок.
Что касается последнего пункта об агрегации Dsup на хроматине, следует отметить, что нет никаких доказательств агрегации Dsup на массивах нуклеосом, как это видно в анализах частичного переваривания MNase (Рисунок 3B и Рисунок 4C), а также в обширный анализ переваривания MNase (рис. 3C) массивов нуклеосом, содержащих Dsup.
6) Hypsibius dujardini, особенно штамм, используемый для геномного / молекулярного анализа, недавно был повторно идентифицирован как новый вид, Hypsibius instanceplaris. Авторы должны использовать новое название вида с кратким объяснением повторной идентификации вида. см.) Gąsiorek P, et al., 2018.
Спасибо за обновление. Мы изменили « H. dujardini » на « H. instanceplaris » во всем тексте и на рисунках и процитировали соответствующую статью (Gąsiorek et al., 2018). Во введении мы включили следующее заявление:
«Примечание: штамм тихоходок, который был обозначен как Hypsibius dujardini в Yoshida et al., 2017, с тех пор оказался новым видом, который теперь получил название Hypsibius instanceplaris (Gąsiorek et al., 2018). В этой статье мы будем использовать новую терминологию «.
https://doi.org/10.7554/eLife.47682.014Этот подраздел Имена и таксономия содержит исчерпывающий список всех названий белка, от широко используемых до устаревших, чтобы можно было однозначно идентифицировать белок. Названия белков i | Рекомендуемое название: Белок-супрессор поврежденийИнформация, подобранная вручную, основанная на утверждениях в научных статьях, не подтвержденных экспериментально. Ручное утверждение, основанное на заключении в i
|
Этот подраздел Имена и таксономия указаны названия генов, которые кодируют последовательность (и) белка, описанную в записи. Четыре существуют разные токены: «Имя», «Синонимы», «Упорядоченные имена локусов» и «Имена ORF». Названия генов i | Название: Dsup Ручное утверждение, основанное на мнении в i
Ручное утверждение, основанное на мнении в i
|
Этот подраздел раздела Имена и таксономия содержит информацию об именах организм, являющийся источником белковой последовательности. Организм i | Ramazzottius varieornatus (Водяной медведь) (Тихоходка) |
В этом подразделе раздела Имена и таксономия показан уникальный идентификатор, присвоенный NCBI организм-источник белка. Это называется таксономическим идентификатором или таксидом. Таксономический идентификатор i | 947166 [NCBI] |
Этот подраздел раздела Имена и таксономия содержит происхождение таксономической иерархической классификации источника. организм. В нем перечислены узлы в том виде, в каком они появляются сверху вниз в таксономическом дереве, с более общей группировкой, указанной первой. Таксономическая линия i | клеточных организмов ›Eukaryota› Opisthokonta ›Metazoa› Eumetazoa ›Bilateria ›Протостомии› Ecdysozoa ›Панартроподы› Tardigrada ›Eutardigrada› Parachela ›Hypsibioidea› Ramazzottiidae ›Ramazzottius |
Этот подраздел раздела Имена и таксономия предназначен для записей, которые являются частью proteome , то есть набора белков, которые, как считается, экспрессируются организмами, чьи геномы полностью секвенированы. Протеомы i |
|
Расчетное структурное исследование на ДНК-защитная роль уникального для тихоходок белка Dsup
Dsup является внутренне неупорядоченным белком, что подтверждается прогнозированием нарушений и структурным моделированием
Выравнивание между Dsup и Dsup-подобными последовательностями, показанное на рис. сегменты, о которых сообщалось ранее: два коротких (11 и 15 аминокислот) идентичных мотива, последовательность NLS из 22 аминокислот, расположенная в С-концевой области, и мотив из 8 аминокислот нуклеосом-связывающего домена HMGN.Аминокислотный состав обеих последовательностей обнаруживает структурный интерес (рис. 1b). В Dsup сумма небольших неполярных (69G + 72A) и полярных (79S + 31T) аминокислот составляет 56,4% от общего количества. Существует большое количество заряженных остатков с преобладанием положительных (59K + 12R) над отрицательными (29D + 19E) аминокислот, которые вместе составляют 26,7% от общего количества и дают общий заряд +23. В Dsup-подобных аминокислотах имеется также большинство небольших неполярных (22G + 76A) остатков, но есть гораздо меньше мелких полярных (26S + 15T) аминокислот, так что все маленькие остатки теперь составляют 42.4% от общей суммы. Сумма заряженных положительных (51K + 11R) и отрицательных (18D + 24E) остатков составляет 31,7% от общего количества с чистым зарядом +20. Общими чертами обеих последовательностей является то, что они (i) не содержат цистеинов, (ii) почти не содержат цистеинов. отсутствие ароматических остатков и (iii) очень мало неполярных аминокислот с длинными боковыми цепями (4I + 1L в Dsup и 5I + 1L в Dsup-подобных). Поскольку цистеины обычно участвуют в образовании дисульфидной связи и неполярные, особенно ароматические аминокислоты способствуют гидрофобной агрегации (объемный триптофан отсутствует в обоих белках), очень низкое количество этих упорядочивающих остатков вместе с большим количеством маленьких и заряженных аминокислот в этих двух белках согласуется с их внутренней неупорядоченной природой, показанной ниже.
Выравнивание на рис. 1a также показывает эквивалентное расположение сегмента, состоящего из 83 аминокислот в Dsup и 42 аминокислот в Dsup-подобных, для которых мы получили структуры, основанные на локальной гомологии (рис. 1c). Несмотря на то, что мы не ожидали получить основанную на гомологии структурную модель всех белков, мы все же попробовали этот подход, чтобы выяснить, возможно ли моделирование гомологии хотя бы для некоторых областей. Swiss-Model предоставила для Dsup две маленькие α-спиральные модели для сегментов последовательностей 136–181 и 185–222 (рис.1c слева), используя в качестве матриц область белка катенина-α-2 мыши (PDB id 4ONS) и область белка аннексина A8 человека (PDB id 1W45), соответственно. Для Dsup-подобного сегмента Swiss-Model предсказала для сегмента 114–155 модель, состоящую из α-спирали и двух коротких спиралей 3 10 (рис. 1c справа), используя в качестве матрицы область бактериальной дегидрогеназы (PDB id 1XDW). Интересно, что Hashimoto et al. сообщили об анализе in silico последовательности Dsup, в котором α-спиральная область была предсказана для сегмента 142-206, тогда как не было предсказано никакой вторичной структуры для оставшейся последовательности (за исключением нескольких крошечных β-цепей) 14, 19 .Хотя эти авторы не касались предсказания беспорядков, они предположили, что Dsup «… может функционировать с гибкой структурой, а не в жесткой форме, например, как физический щит ДНК» (стр. 6 в Ref. 19 ).
Мы использовали несколько предикторов беспорядка, и их результаты (рис. 1d) вполне убедительны: за исключением области последовательности ~ 140–220, три из четырех методов предсказали для Dsup неупорядоченную структуру, а оставшийся метод, IUPred2, предсказал в этой область с более низкой оценкой беспорядка (область, заштрихованная светло-зеленым на рис.1d слева). Большинство предикторов расстройства основаны на знаниях. PONDR — это нейронная сеть, обученная на известных неупорядоченных последовательностях, и связанный с ней алгоритм VLXT включает информацию из неупорядоченных областей, охарактеризованных рентгеновскими лучами. DisEMBL и DISOPRED3 обучаются на структурных данных из известных неупорядоченных областей, таких как неназначенные электронные плотности, определенные как Замечание 465 в файлах структуры или петли / катушки с высокими тепловыми факторами. Помимо структурной информации, DISOPRED3 также использует данные о последовательностях, о которых известно, что они неупорядочены, хотя они не имеют структуры.Напротив, IUPred2 предсказывает беспорядок, вычисляя энергии взаимодействия между всеми парами аминокислот в последовательности. Его основное предположение состоит в том, что неупорядоченные белки имеют аминокислотный состав, который исключает благоприятные взаимодействия между остатками для стабилизации четко определенной структуры. IUPred2 использует быстрый и надежный статистический потенциал, разработанный для вычисления энергий взаимодействия пар аминокислот, не полагаясь на базы данных, и, таким образом, методологически отличается от любого другого предсказателя расстройства.Несмотря на различные подходы, лежащие в основе этих предикторов беспорядка и предиктора вторичной структуры, использованного в [3]. 19 для Dsup, все они замечательно согласуются с результатами нашего моделирования. Эти находки можно резюмировать, утверждая, что Dsup является внутренне неупорядоченным белком (IDP) и что возможно, что динамические конформационные изменения, которые характеризуют IDPs, д. Обнаруживать временные α-спиральные локальные сегменты в области 140-220. Аналогичные результаты были получены для Dsup-подобного: беспорядок предсказывается для большей части последовательности, за исключением сегмента ~ 115–155 (заштрихованный светло-зеленым на рис.1d справа), для которого три из четырех методов снова не предсказывают расстройства, а IUPred2 предсказывает более низкий балл расстройства. Кроме того, в двух небольших областях ~ 50–70 и ~ 175–210 теперь двумя методами предсказывается отсутствие беспорядка. Что касается сегментов последовательности, строго консервативных в Dsup и Dsup-подобных (рис. 1a; области, заштрихованные светло-серым цветом на рис. 1d), все методы согласуются в предсказании немного более высокого беспорядка для второго мотива (262–275), чем для первого. один (90–100), и оба они обрамляют центральную область (заштрихованы светло-зеленым на рис.1г).
В соответствии с предсказаниями беспорядков, структуры 3D-модели, полученные с помощью I-TASSER для полных последовательностей (рис. 1e), фактически предсказывают, что оба белка являются IDP. Учитывая, что IDP существуют как динамический ансамбль конформаций, которые в значительной степени меняются со временем, без единой равновесной структуры, эти модели I-TASSER следует рассматривать только как одну из возможных конформаций. Как показано ниже для Dsup, именно динамическая эволюция структуры обеспечивает важную информацию о ее поведении в отношении взаимодействия с ДНК.Неупорядоченная структура Dsup показывает заряженные аминокислоты, сгруппированные в отдельные участки (рис. 1e слева). В то время как пять из них также имеют кислотные остатки, две области имеют исключительно основные остатки: сегмент 260–283 (4K + 3R) и С-конец (14K + 3R), который имеет положительный заряд +17. Как обсуждается ниже, он является ли этот последний положительный сегмент тем, который преимущественно движется в сторону ДНК, что согласуется с гипотезой Hashimoto et al. что C-конец Dsup и Dsup-подобных белков будет участвовать в ассоциации ДНК 14, 19 Dsup-подобная модель также показывает заряженные остатки, сгруппированные по участкам (рис.1д справа). Теперь существует единственный регион, содержащий только основные остатки: сегмент 210–230 (7K + 3R). Напротив, C-концевой участок (15K + 8R) также имеет шесть кислотных остатков, что дает локальный положительный заряд + 17, такой же, как у Dsup.
Обнаружено, что положительные и отрицательные заряды разбросаны по всем структурам как Dsup, так и Dsup-подобных, за исключением аккуратно разделенных областей с накоплением положительных остатков (без учета этих областей чистый заряд составляет всего -1 в обоих белках).Это открытие можно рационализировать, предположив, что постоянное движение неупорядоченной цепи допускает баланс зарядов в качестве общего стабилизирующего фактора, в то время как существование разделенных областей с подавляющим преобладанием положительных зарядов предполагает, что для стабилизации необходим внешний фактор, такой как соединение с ДНК. их. Стоит отметить, что кроме С-концов, сегмент с исключительно основными остатками (отмечен оранжевыми овалами на рис. 1д) содержит второй из двух консервативных мотивов между белками.Хотя для этих мотивов еще не предложена функция, это открытие, предоставленное структурными моделями, может указывать на предполагаемую роль, которая представлена ниже после обсуждения результатов моделирования MD.
Известно, что экспрессия генов, кодирующих специфичные для тихоходок IDP, повышается в ответ на сушку. 28 . Большое количество этих IDP было идентифицировано у H. instanceplaris , и способность 11 цитозольных обильно растворимых при нагревании белков витрифицироваться была продемонстрирована in vitro и при экспрессии в дрожжах 28 .Было высказано предположение, что если другие устойчивые к высыханию организмы полагаются на трегалозу для адаптации к высыханию, тихоходки будут полагаться на этих ВПЛ, чтобы пережить почти полное высыхание 28 . Эти находки согласуются с описанными свойствами белков позднего эмбриогенеза изобилия (LEA), других средств защиты от высыхания, которые используют свои неупорядоченные гибкие структуры для защиты других молекул путем экранирования их или для образования материала, поддерживающего стеклование во время высыхания 29 .Следовательно, предположение о том, что неупорядоченный белок может функционировать как гибкая структура, защищающая ДНК от повреждений, вызванных облучением, кажется разумным и действительно демонстрируется ниже. Неожиданно низкое сходство последовательностей между Dsup из R. varieornatus и Dsup-like из H. instanceplaris , таким образом, может указывать на то, что они находились под слабым селективным давлением во время эволюции. Это известная особенность IDP, поскольку аминокислоты в неупорядоченных областях могут изменяться без физических ограничений для поддержания определенной структуры 30,31,32 .Подводя итог этому разделу, Dsup и Dsup-подобные имеют: (i) схожий профиль IDP, (ii) центральную область с потенциальной вторичной структурой в виде спиральных мотивов, фланкируемых двумя короткими сегментами последовательности, строго консервативными, (iii) предсказанная трехмерная модель, которая показывает аналогичную неупорядоченную структуру с кластерами заряженных аминокислот, (iv) отдельный C-концевой кластер с накоплением положительно заряженных остатков, который включает консервативную последовательность NLS, (v) аналогичный кластер положительных аминокислот, который включает второй из двух строго консервативных мотивов последовательности, (vi) подобный чистый положительный заряд, (vii) идентичный чистый заряд — 1, полученный после исключения кластеров положительных зарядов.Вместе с ранее идентифицированными консервативными сегментами последовательности наши данные убедительно свидетельствуют о том, что Dsup и Dsup-подобные могут иметь общую функцию, в которой первостепенное значение имеет нарушение.
Начальная геометрия комплексов Dsup-ДНК демонстрирует значительную электростатическую комплементарность
С целью изучения динамического поведения Dsup мы выполнили МД-моделирование изолированного белка и двух комплексов белок-ДНК со стехиометрией 1: 1 и 2: 1 . Размер сольватационного бокса, необходимый для обращения к этим системам с помощью расчетов МД для всех атомов, дает очень большое количество атомов: 414 174 в комплексе 1: 1 и 755 424 в комплексе 2: 1, из которых только 9 561 и 15 503 атома. соответственно соответствуют белку и ДНК.В случае 1: 1 мы устанавливаем начальную геометрию комплекса (рис. 2а), помещая одну молекулу Dsup так, чтобы она могла взаимодействовать почти со всей идеальной моделью ДНК (см. «Методы»). Поскольку экспериментальные данные показывают, что Dsup взаимодействует со свободной ДНК 14, 19 и с нуклеосомами 25 независимо от нуклеотидной последовательности, мы смоделировали ДНК в наиболее часто встречающейся конформации B на основе случайно сгенерированной 57-нуклеотидной последовательности. Учитывая, что динамика, полученная с помощью МД-расчетов, изменяет геометрию как белка, так и ДНК из-за взаимодействия между ними и с водой, эта идеальная структура ДНК является лишь отправной точкой для динамического исследования.Первоначальное разделение между Dsup и ДНК было достаточно большим, чтобы позволить моделированию MD сблизить обе молекулы, но не достаточно далеко, чтобы потребовалось чрезмерное количество шагов моделирования только для того, чтобы поместить две молекулы на расстояние взаимодействия (рис. 2а). В случае 2: 1 мы добавили к этой геометрии 1: 1 вторую молекулу Dsup на немного большем расстоянии от ДНК и менее параллельную ориентацию по отношению к ДНК (рис. 2b), чтобы позволить моделирование МД перемещать вторую молекулу Dsup. под действием как ДНК, так и первой молекулы Dsup (см. расстояния на рис.2).
Рисунок 2Структура и электростатический потенциал Пуассона-Больцмана исходной геометрии комплексов Dsup-ДНК. (a ) Виды сбоку (слева) и сверху (справа) исходной геометрии комплекса Dsup-ДНК. Белок показан в виде зеленой ленты с областями, соответствующими сегментам консервативной последовательности, окрашенным, как на фиг. 1а для Rv Dsup. ДНК показана в виде шариков и палочек с атомами P оранжевым, атомами O красным, атомами N синим и атомами C голубым. Минимальные расстояния между ДНК и тремя сегментами Dsup, расположенными ближе к ДНК, указаны на виде сверху.( b ) Те же виды для комплекса (Dsup) 2 -ДНК. Первая молекула Dsup, названная DsupA, показана как ( a ), а вторая молекула, названная DsupB, показана в виде синей ленты с той же окраской сегментов последовательности, что и на фиг. 1a для Rv Dsup. Минимальные расстояния между ДНК и тремя сегментами второй молекулы Dsup, ближайшими к ДНК, указаны на виде сверху. ( c ) Вид сбоку и сверху PB-EP, картированного на поверхности белка и ДНК в комплексе Dsup-ДНК.( d ) Те же виды, что и в ( c ) для комплекса (Dsup) 2 -ДНК. Полоса представляет собой цветовой код для диапазона значений PB-EP (в единицах kT, / и ) в этих сопоставлениях. ( e ) Изоповерхности PB-EP + 0,1 (светло-синяя сетка) и -0,1 (светло-красная сетка) наложены на поверхности, показанные в (c ) для комплекса Dsup-ДНК. ( f ) Те же виды для комплекса (Dsup) 2 -ДНК.
Картирование PB-EP на поверхности Dsup и ДНК показывает общую электростатическую комплементарность как в соотношении 1: 1 (рис.2в) и комплексов 2: 1 (рис. 2г). Заметно отрицательный EP на поверхности ДНК, в котором преобладают отрицательные фосфатные группы в 57 нуклеотидах каждой цепи, контрастирует с преобладающим положительным EP на поверхности Dsup. Пространственные домены, определяемые изоповерхностями PB-EP -0,1 и + 0,1 в этих начальных геометриях (рис. 2e, f), показывают, что отрицательная изоповерхность, равномерно расположенная вокруг ДНК, дополняет большие участки положительных изоповерхностей, которые возникают из белков. Как показано ниже, динамическая эволюция этих систем сближает Dsup и ДНК за счет усиления этой электростатической комплементарности.
Электрическое поле, создаваемое молекулярным EP V ( r ), представляет собой отрицательный градиент потенциала, E = −∇ V ( r ). Линии поля E показаны на рис. 3a, b для исходной геометрии двух комплексов Dsup-ДНК, показанных на рис. 2. Дополнительные линии поля показаны на рис. 3c, d для двух геометрий, полученных при перемещении. белки дальше от ДНК за счет горизонтальной трансляции в ориентациях, показанных для двух комплексов на рис.3а, б. Линии электрического поля начинаются с положительных зарядов и заканчиваются отрицательными зарядами, которые представлены на рис. 3 синим и красным цветами соответственно. В ( r ) выражается в единицах kT / e ( k — постоянная Больцмана, T — абсолютная температура и e — заряд электрона), так что при комнатной температуре 1 кТл / e = 0,0258 Дж / Кл = 0,0258 В. Поскольку электрическое поле имеет единицы вольт / длину, одна кТл -эквивалентная единица электрического поля E в молекулярных системах равна 0.0258 В / Å = 2,58 × 10 8 В / м. Сила (то есть величина) электрического поля в любой области пространства пропорциональна плотности силовых линий в этой области. Следовательно, электрическое поле, показанное на рис. 3 для исходной геометрии, в которой Dsup и ДНК разделены на значительное расстояние, указывает на то, что большое количество зарядов в обеих молекулах создает сильные электрические поля. Даже при больших межмолекулярных расстояниях на рис. 3а, б эти поля составляют ~ 0,15 В / Å = 1,5 · 10 9 В / м и ~ 0.08 В / Å = 8 × 10 8 В / м (рис. 3в, г). Для сравнения сообщалось, что внешние электрические поля силой ~ 10 9 В / м ускоряют фотосинтетические реакции на порядок, и что внутренние поля также примерно 10 9 В / м генерируются на активных участках. ферментов 33 .
Рисунок 3Электрическое поле E в начальной геометрии комплексов Dsup-ДНК. ( a ) Виды линий поля сбоку (слева) и сверху (справа) для E = 6 kT -эквивалентных единиц в исходной геометрии комплекса Dsup-ДНК, показанного на рис.2а. ( b ) Те же виды, что и использованные в ( a ) для комплекса (Dsup) 2 -ДНК. ( c ) Вид сбоку линий поля для E = 3 kT -эквивалентных единиц в геометрии комплекса Dsup-ДНК в результате перемещения Dsup дальше от ДНК в геометрии, показанной в ( a ). ( d ) Вид сбоку силовых линий для E = 3 kT -эквивалентных единиц в геометрии (Dsup) 2 -ДНК-комплекса в результате перемещения обеих молекул Dsup дальше от ДНК в геометрия показана в ( b ).Линии поля на изображениях ( a ) — ( d ) начинаются на (+) зарядах и заканчиваются (-) зарядами, обозначенными синим и красным цветами соответственно.
Полноатомное моделирование МД показывает отличное локальное поведение областей Dsup и динамическую роль С-концевого домена в присутствии ДНК
В этом и следующих разделах мы представляем результаты моделирования МД всех атомов за 100 нс. для следующих систем: изолированный Dsup (две траектории), Dsup-ДНК 1: 1 комплекс с (i) полной цепью Dsup, (ii) цепью, лишенной C-концевого домена, и (iii) другой цепью, состоящей только из этого домен и (Dsup) 2 -ДНК 2: 1 комплекс.В комплексе 1: 1 начальные геометрии (ii) и (iii) были таковыми из соответствующих сегментов в начальной геометрии полного Dsup. В комплексе 2: 1 две молекулы Dsup далее обозначаются как DsupA и DsupB (см. Фиг. 2). Поскольку, как уже упоминалось, Dsup, по прогнозам, внутренне неупорядочен, за исключением, возможно, средней области, охватывающей приблизительно остатки 140-220, большие динамические изменения ожидаются вдоль его основной цепи, за исключением, возможно, этой средней области. Однако очень маловероятно, что большие элементы вторичной структуры могут появиться в наших МД-симуляциях, потому что локальные процессы сворачивания, которые будут задействованы, происходят в гораздо более длительное время, чем те, которые поддаются МД-исследованиям с очень большими белками, такими как Dsup.Тем не менее, во время постоянных структурных изменений полного IDP ожидается, что (а) временные крошечные элементы вторичной структуры появятся в небольших разреженных сегментах, и (б) разные области скелета будут демонстрировать различную подвижность.
Результаты расчета среднеквадратичных отклонений (RMSD) с атомами основной цепи по всем атомным 100 нс МД моделирования показаны на рис. 4a, сначала как график для всей магистрали, а затем как набор графиков тепловой карты, отображающих значения из 445 остатков.Неупорядоченное движение белка в целом отражается в высоком среднеквадратичном отклонении полной цепи (первая панель на рис. 4a), которая показывает очень похожую картину во всех системах. Подвижность, измеренная с помощью RMSD, быстро увеличивается в течение первых 10 нс, затем увеличивается более плавно в течение следующих 20 нс и на ~ 40 нс стабилизируется на большом значении RMSD ~ 30 Å на всех траекториях. Этот результат предполагает последовательное динамическое поведение неупорядоченной структуры нашей модели белка Dsup.Графики тепловой карты, показывающие среднеквадратичное отклонение всех остатков по результатам моделирования, показывают четкую закономерность для разных сегментов последовательности. Общей чертой всех траекторий является большая подвижность некоторых участков последовательности, таких как первые 50 N-концевых аминокислот, область вокруг остатка 300 и большая часть C-концевых остатков за пределами положения ~ 350, которая включает обе нуклеосомы. -связывающая последовательность белков HMGN (363–370) и последовательность NLS (383–404). Первый сегмент 90–100, строго консервативный в Dsup и Dsup-like, обнаруживает низкую подвижность на большинстве траекторий, тогда как второй строго консервативный сегмент 262–275 обнаруживает более высокую подвижность в комплексах ДНК, чем в двух изолированных траекториях.Стоит отметить, что эти особенности находятся в разумном согласии с (а) предсказанием беспорядка на рис. 1d, где видно, что два консервативных сегмента действительно имеют разные оценки беспорядка в большинстве методов, и (b) 3D-модели на рис. .1e, где обнаружено, что второй консервативный сегмент является частью положительно заряженного кластера, который может участвовать во взаимодействиях ДНК. Что касается центральной области, для которой были получены модели на основе локальной гомологии (136–222), все траектории, кроме траектории DsupA в комплексе 2: 1, показывают меньшие значения RMSD в этой области последовательности ~ 150–250, чем в большинстве траекторий. оставшиеся остатки.Эти графики предполагают, что две молекулы Dsup в комплексе 2: 1 влияют друг на друга, поскольку они демонстрируют качественно различное поведение в этой средней области.
Рис. 4Структурные данные, рассчитанные для Dsup вдоль полностью атомных MD траекторий 100 нс в исследованных системах. ( a ) График значений RMSD для белка Dsup (первая панель) и графики RMSD для 445 остатков, вычисленных с атомами основной цепи. ( b ) График RMSF. Области последовательностей, заштрихованные разными оттенками, соответствуют сегментам последовательности, указанным на рис.1г такими же цветами. ( c ) Радиус вращения.
Последний результат тем более очевиден в пространственных колебаниях остатков, измеренных с помощью среднеквадратичного колебания (RMSF), показанного на рис. 4b. Фактически, молекулы DsupA и DsupB в комплексе 2: 1 демонстрируют заметно различное поведение, особенно в области ~ 150–250, в которой DsupA имеет гораздо большие флуктуации, чем DsupB. Dsup в комплексе 1: 1 показывает в целом меньшую флуктуацию, чем в остальных системах, результат, который можно объяснить близостью между большими структурными сегментами белка и ДНК, которые появляются на ранних этапах динамики, предсказанной расчетами МД, как обсуждалось. ниже.
Даже несмотря на то, что радиус инерции (R G ) не может считаться характерным свойством в IDP, стоит отметить его тенденции в системах Dsup. Во время последнего моделирования R G демонстрирует тенденцию к снижению в трех белках Dsup в комплексе с ДНК, тогда как он стабилизируется или даже увеличивается на двух траекториях изолированного Dsup (фиг. 4c). Кроме того, значения R G в двух из трех случаев, в которых Dsup взаимодействует с ДНК, составляют примерно половину значений в двух изолированных случаях Dsup.Этот результат предполагает склонность к более компактной структуре в присутствии ДНК.
В предыдущих экспериментальных отчетах было показано, что C-концевой домен Dsup необходим и достаточен для связывания ДНК 14, 19 . Стремясь изучить динамическую роль этого домена во взаимодействии с ДНК, мы выполнили дополнительные МД-расчеты в комплексе 1: 1 для цепи Dsup 1–370, не содержащей C-концевой домен, и для другой цепи 370–445, состоящей из этого домена. один. Сравнение RMSD, R G и RMSF для этих цепей и для соответствующих сегментов в полной цепи, все в комплексе с ДНК, показано на рис.5. Основные результаты следующие. (i) RMSD-значения цепочки 1–370 очень похожи на значения RMSD для всей цепочки, но становятся меньше от ~ 40 нс до конца моделирования, демонстрируя аналогичный профиль (рис. 5a). Как и ожидалось, исходя из их относительной длины, значения R G в цепях 1-370 меньше, чем у полного белка, с аналогичным профилем на протяжении всего моделирования (Рис. 5b). (ii) Значения RMSD и R G цепи 370–445 (только C-концевой домен), очевидно, намного меньше, чем у двух других цепей, но теперь есть драматические изменения.С начального времени до ~ 40 нс, а затем от 60 нс до конца моделирования и RMSD, и R G показывают относительно плоский профиль. Однако между ~ 40 и 60 нс у изолированного С-концевого домена RMSD даже выше, чем у цепей 1-370 (рис. 5a), и значения R G аналогичны (рис. 5b), результаты, которые на первый взгляд могут быть кажутся удивительными, если учесть, что при сравнении учитываются разные длины цепей. (iii) RMSD C-концевого домена в полной цепи также показывает заметное увеличение на ~ 40 нс, остается на больших значениях до ~ 80 нс, а затем уменьшается, становясь подобным таковому для полного белка (рис.5а). R G этого сегмента в полной цепочке демонстрирует ровный плоский профиль, из которого видно, что его размер практически не изменился (рис. 5b). (iv) Сегмент 1–370 в полном белке следует заметно схожей тенденции в значениях RMSD и R G с таковым для полного белка, тогда как C-концевой домен в белке отвечает за заметно большие колебания RMSD от ~ 40 нс до ~ 80 нс (рис. 5а) и другая тенденция R G , особенно до ~ 40 нс (рис.5б). (v) Основа цепочки 1–370 показывает общие более низкие значения RMSF, чем полная цепочка, и не имеет больших колебаний, которые полный Dsup показывает в нескольких сегментах (~ 50–90, ~ 125–135, ~ 195–215 и ~ 295– 310). Остов изолированного C-концевого домена обнаруживает заметно меньшие колебания, чем у полного белка в этом домене, что особенно заметно в сегменте ~ 405-430 сразу после последовательности NLS (Fig. 5c).
Рис. 5Структурные данные, вычисленные при моделировании всего атома МД за 100 нс для различных цепей Dsup в комплексе Dsup-ДНК 1: 1.( a ) График сравнения RMSD, вычисленного с атомами остова для (i) полной цепи 1–445, (ii) для цепи 1–370 без С-концевого домена, (iii) для цепи 370–445, состоящей только из этого C-концевого домена и (iv, v) для двух соответствующих сегментов в полной цепи Dsup. ( b ) График сравнения значений радиуса инерции для цепей (i) — (v). (c ) RMSF для цепей (i) — (iii). Области последовательностей, заштрихованные разными оттенками, соответствуют сегментам последовательности, указанным на рис.1г такими же цветами.
Чтобы понять результаты динамического поведения изолированного C-концевого домена, необходимо принять во внимание, что его исходная геометрия была взята из соответствующего сегмента 370–445 в модельной структуре полного Dsup, который представляет собой компактный положительно заряженный кластер (382–445 на рис. 1д). Этот выбор был сделан для того, чтобы предоставить дополнительные ключи к разгадке динамики взаимодействия С-концевого домена с ДНК, поскольку рис. 5 предполагает, что этот домен, в частности, имеет большую подвижность, чем остальная часть цепи в течение большей части времени моделирования.MD-траектория цепи 370–445 с одним только C-концевым доменом выявляет баланс между ее динамическим движением и зарядовым взаимодействием с ДНК. Эта траектория проиллюстрирована снимками на вспомогательном рис. 1: в дальнейшем время относится к этим снимкам. Домен начинает движение в виде компактной цепочки (низкий R G ) с низкой подвижностью (низкий RMSD), поскольку его исходная геометрия представляет собой компактный кластер (0–8 нс). Когда он все еще находится далеко от ДНК, отталкивание между его положительными зарядами начинает расширять его основу (увеличивая R G ), что, в свою очередь, вызывает большую подвижность (увеличение RMSD) в воде (16–32 нс).Пока эти эффекты продолжают растягивать цепь (наибольшее RMSD и R G ), один из ее концов приближается к ДНК (40–64 нс). Когда притяжение между положительными зарядами С-концевой цепи и отрицательными зарядами ДНК достаточно велико из-за их более близкого соседства, основная цепь сжимается (низкий R G ), уменьшая, таким образом, ее подвижность (низкий RMSD), пока она не плотно прикрепляется к ДНК (72–100 нс). Эта динамика, при которой изолированный С-концевой домен упаковывается в течение значительной части времени моделирования, вызывает более низкую флуктуацию (RMSF) только этого сегмента 370–445, чем флуктуацию соответствующего сегмента в полном белке, в котором он является частью ВПЛ, что сильно влияет на его мобильность (рис.5в).
Динамические изменения расстояний Dsup-ДНК дают представление о взаимодействии
Взаимодействие между Dsup и ДНК теперь анализируется с помощью минимальных (рис. 6) и максимальных (рис. 7) расстояний между обеими молекулами, вычисленных по МД моделирования. Эти расстояния были рассчитаны для каждой рамки траектории как минимальное и максимальное значения среди всех расстояний между атомами Cα во всех остатках белка и атомами P во всех нуклеотидах в двух цепях ДНК.Минимальное расстояние для белка в целом (рис. 6а) начинает уменьшаться в самом начале моделирования, а затем стабилизируется на небольшом значении на 9 нс в комплексе 1: 1 и на 40 нс в комплексе 2: 1. . Поскольку исходная геометрия комплекса 2: 1 была получена путем добавления второй молекулы Dsup (DsupB) к исходной геометрии комплекса 1: 1, начальные значения этого минимального расстояния одинаковы для комплекса 1: 1 и для DsupA в комплексе 2: 1. Однако, хотя две молекулы Dsup в комплексе 2: 1 достигают постоянного минимального расстояния до ДНК практически одновременно, им требуется гораздо больше времени, чем Dsup в комплексе 1: 1.Этот результат согласуется с упомянутым выше взаимным влиянием обеих молекул Dsup в комплексе 2: 1. Ближайшее расстояние между Dsup и ДНК во всех случаях составляет примерно 4 Å (рис. 6а), что свидетельствует о тесном и стабильном взаимодействии. Сравнение минимальных расстояний полного белка с таковыми для цепи, лишенной C-концевого домена, и с расстояниями только этого домена обеспечивает дальнейшее понимание роли C-концевой области во взаимодействии с ДНК (рис. 6b). В отсутствие C-концевого домена цепь из 370 остатков впервые достигает минимального значения за 30 нс, но почти сразу же снова увеличивается, и только через 50 нс восстанавливаются минимальные значения, хотя с небольшими увеличениями, которые даже больше в конце моделирования (рис.6б). Большая подвижность изолированного С-концевого домена, обсуждавшаяся выше, отражается в больших колебаниях его минимального расстояния до ДНК до уменьшения на 40 нс, за которым следует большое увеличение между 40 и 60 нс, чтобы наконец достичь устойчивого минимального расстояния ~ 4 Å. (Рис. 6б).
Рис. 6Минимальные расстояния между Dsup и ДНК, вычисленные при моделировании всего атомного MD 100 нс для комплексов Dsup-ДНК. Это минимальные расстояния между любым атомом Cα Dsup и любым атомом P ДНК.( a ) Минимальные расстояния, вычисленные с полными цепочками Dsup в двух комплексах. ( b ) Минимальные расстояния в комплексе 1: 1, рассчитанные с использованием полной цепи Dsup, с цепью, лишенной C-концевого домена 370–445, и с цепью, состоящей только из этого домена. ( c ) График тепловой карты минимальных расстояний остатков для полной цепи Dsup в комплексе 1: 1. ( d ) График тепловой карты для цепи 1–370 без С-концевого домена (нижняя часть) и для цепи 370–445, состоящей только из этого домена (верхняя часть).( и ) Графики тепловой карты для молекул DsupA и DsupB в комплексе 2: 1.
Рис. 7Максимальные расстояния между Dsup и ДНК, вычисленные при моделировании всего атомного MD 100 нс для комплексов Dsup-ДНК. Это максимальные расстояния между любым атомом Cα Dsup и любым атомом P ДНК. ( a ) Максимальные расстояния, вычисленные с полными цепочками Dsup в двух комплексах. ( b ) Максимальные расстояния в комплексе 1: 1, рассчитанные с использованием полной цепи Dsup, с цепью, лишенной C-концевого домена 370–445, и с цепью, состоящей только из этого домена.( c ) График тепловой карты максимальных расстояний остатков для полной цепи Dsup в комплексе 1: 1. ( d ) График тепловой карты для цепи 1–370 без С-концевого домена (нижняя часть) и для цепи 370–445, состоящей только из этого домена (верхняя часть). ( и ) Графики тепловой карты для молекул DsupA и DsupB в комплексе 2: 1.
Графики тепловой карты на рис. 6 дают минимальные расстояния от всех остатков Dsup до ДНК по результатам моделирования MD. Замечено, что приблизительно сто С-концевых остатков неизменно достигают наибольшей близости к ДНК при времени моделирования ~ 40 нс во всех случаях.Однако остальная часть цепочки Dsup ведет себя иначе. В комплексе 1: 1 более низкие значения минимальных расстояний до ДНК также появляются во второй половине моделирования в других областях полной цепи (рис. 6c), таких как N-концевой сегмент длиной примерно в сотню остатков и большие области. между положениями ~ 250 и ~ 350. Все эти области также достигают наибольшего сближения с ДНК примерно за 40 нс. По истечении этого времени минимальные расстояния, превышающие 50 Å, не обнаруживаются на рис. 6c, что свидетельствует о тесном межмолекулярном взаимодействии между Dsup и ДНК (см. Вспомогательное видео 1).Роль С-концевого домена в близости к ДНК наглядно демонстрируется при сравнении этого рисунка 6c с рисунком 6d, оба относятся к одному и тому же комплексу 1: 1. Изолированный C-концевой домен (верхняя часть графика d) показывает картину, очень похожую на структуру двух молекул Dsup в комплексе 2: 1 (рис. 6e), но не на картину полного Dsup (рис. 6c): в середине значения> 50 Å минимальных расстояний в течение первых 40 нс, а затем низкие значения <15 Å в течение оставшихся 60 нс (рис. 6d). Опять же, цепь без С-концевого домена (нижняя часть графика d) демонстрирует общую картину, довольно отличную от той, которая наблюдается в соответствующем сегменте 1–370 полного белка (рис.6c) и более сходен с таковой для обеих молекул Dsup в комплексе 2: 1 (рис. 6e). Распределение минимальных расстояний в этом комплексе 2: 1 показывает, что наиболее близкое приближение к ДНК теперь появляется исключительно в С-концевой области двух молекул Dsup (Рис. 6e). Если посмотреть на их относительное расположение по отношению к ДНК в начальной геометрии МД-моделирования (см. Рис. 2b), эти тепловые карты предполагают, что каждая молекула Dsup движется по направлению к ДНК в течение первых 40 нс, все еще находясь вдали от ДНК и затем ориентировать их С-концы ближе к ДНК, таким образом оставаясь в течение следующих 60 нс (см. вспомогательное видео 2).Этот результат согласуется с неоднократными наблюдениями за изменениями, происходящими в это время, которые обсуждались выше для рис. 6а. Наконец, сегмент 262–275 (второй из двух строго консервативных мотивов в Dsup и Dsup-подобных) близок к ДНК на всех графиках тепловой карты на рис. 6, что соответствует положению внутри кластера положительно заряженных аминокислот ( 260–283 на рис. 1д, слева).
Максимальные расстояния Dsup-ДНК дают дополнительную картину (рис. 7). Значения, рассчитанные для полных белков (рис.7а) остаются с небольшими изменениями в течение первых 20 нс в обоих комплексах. Но в то время как эти расстояния плавно уменьшаются после этого времени аналогичным образом для Dsup в комплексе 1: 1 и для DsupB в комплексе 2: 1, они увеличиваются и остаются на больших значениях для DsupA в последнем комплексе. Во всех случаях эти расстояния сходятся к общему значению в конце моделирования. Сравнение максимальных расстояний полного Dsup с расстояниями цепи 1–370, лишенной С-концевого домена, и расстояний только этого домена предполагает, что влияние этой С-концевой области на максимальное отделение от ДНК меньше, чем на минимальные расстояния. к ДНК (рис.7б). Это ожидается, поскольку при максимальных расстояниях будут выявлены области, которые специфически отделяются или косвенно остаются отдельно от ДНК, а результаты МД, обсужденные выше, демонстрируют, что C-концевой домен представляет собой область Dsup, которая прикрепляется к ДНК во всех случаях. Цепь без С-концевого домена демонстрирует некоторые флуктуации максимальных расстояний в течение первых 40 нс, что означает, что в отсутствие этого домена подвижность неупорядоченной цепи увеличивается в течение этого времени. Однако эти значения меньше, чем у полных цепочек, и начиная с 40 нс все расстояния сходятся к одному и тому же максимальному значению около 160 Å (рис.7а, б).
Графики тепловой карты на рис. 7, показывающие максимальные расстояния от ДНК всех остатков Dsup, показывают, что большие значения на большей части моделирования соответствуют во всех случаях нескольким сегментам в 150 N-концевых остатках. В комплексе 1: 1 меньшие значения появляются в большой области между положениями 150 и 350 (рис. 7c). Как и предполагалось на рис. 7b, роль C-концевого домена в отделении от ДНК (рис. 7d) слабее, чем роль, которую играет близость к ДНК (рис.6г). Главный эффект теперь состоит в том, что отсутствие этого домена (нижняя часть рис. 7d) уступает место немного большим максимальным расстояниям, чем в полной цепочке в области 150–350 (рис. 7c). Напротив, значения для изолированного C-концевого домена (верхняя часть рис. 7d) довольно похожи на значения, наблюдаемые для полной цепи (рис. 7c). Интересно, что большие различия обнаружены для двух молекул Dsup в комплексе 2: 1 (рис. 7e). В то время как DsupB не показывает особенно больших максимальных значений во времени, DsupA отображает в течение большей части моделирования большой сегмент, охватывающий первую половину последовательности, который показывает значения, значительно превышающие другую половину.Этот результат предполагает, что в присутствии DsupB N-концевая половина неупорядоченной цепи DsupA отодвигается от DsupB как можно дальше, усиливая взаимодействие с ДНК его другой половины, снова показывая, что две молекулы Dsup, взаимодействующие с ДНК, могут влияют друг на друга в отношении своего динамического поведения.
Подводя итог этому разделу, можно сказать, что динамическая эволюция различных частей Dsup во взаимодействии с ДНК демонстрирует различное поведение в двух проанализированных комплексах.В комплексе 1: 1 большая часть Dsup показывает небольшие значения как минимального, так и максимального расстояния до ДНК, предполагая, что большая часть белка тесно взаимодействует с ДНК. Роль C-концевого домена, изучаемого в этом комплексе, кажется первостепенной для того, чтобы позволить полному IDP приблизиться к ДНК. В комплексе 2: 1 присутствие дополнительной молекулы Dsup оказывает влияние на другую молекулу Dsup, которое было выше ожидаемого на основе тепловых карт RMSD (фиг. 4a). Фактически, качественно различное поведение в N-концевой и средней областях, заметное на тепловых картах расстояний (рис.6e, 7e) показывает, что DsupA и DsupB влияют друг на друга. Следует отметить, что на основании анализа последовательности in silico, Hashimoto et al. предположили, что N-концевые и средние области Dsup могут играть роль в ослаблении возможных побочных эффектов, вызванных образованием больших комплексов Dsup-ДНК 14, 19 . Это предположение находится в замечательном согласии с нашими выводами, полученными из анализа динамических изменений расстояний белок-ДНК, происходящих во время моделирования МД.Наконец, что касается изменений расстояний до ДНК, второй из двух мотивов последовательности, строго консервативных в Dsup и Dsup-like (262–275), ближе к ДНК, чем первый мотив (90–100), и не имеет каких-либо особенностей. замечены в последовательности нуклеосом-связывающего домена белков HMGN.
Окончательная геометрия комплексов Dsup-ДНК после 100 нс МД-моделирования показывает, что обе молекулы плотно агрегированы
Отсутствие жесткой структуры и высокая степень динамики наделяют IDP гибкостью для адаптации своей структуры к партнерам по связыванию. дело ДНК.Кроме того, наличие дальнего притяжения между IDP и его партнером способствует образованию так называемых неупорядоченных комплексов. В этих комплексах часто обнаруживается, что области, фланкирующие интерфейс взаимодействия, но не сам интерфейс, остаются неупорядоченными 34 . Окончательная геометрия двух агрегатов Dsup-ДНК, изученных в этой работе (рис. 8 и дополнительный рис. 2), демонстрирует особенности, напоминающие такие неупорядоченные комплексы. Хотя внутреннее расстройство Dsup не позволяет получить четкую, уникальную картину его взаимодействия с ДНК, эти окончательные геометрические формы предоставляют некоторые структурные детали, представляющие интерес.
Рис. 8Конечная геометрия комплексов Dsup-ДНК после моделирования многоатомной MD 100 нс и структурного нарушения в ДНК-связывающих белках. (a ) Комплекс Dsup-ДНК. Области консервативной последовательности отмечены на белковой ленте теми же цветами, что и на рис. 1а. ( b ) Два вида электрического поля E = 7 kT -эквивалентных единиц в этой окончательной геометрии. ( c ) (Dsup) 2 -ДНК комплекс. Области консервативной последовательности отмечены на обеих белковых ленточках (зеленый: DsupA, синий: DsupB) теми же цветами, что и на фиг.1а. ( d ) Два вида электрического поля E = 7 kT -эквивалентных единиц в этой окончательной геометрии. ( e ) Неупорядоченные области (непрозрачные сегменты петли) фактора транскрипции Ets-1 (красная лента) и его спаренный домен белка бокса PAX5 (синяя лента) увеличивают аффинность связывания ДНК почти в 1000 раз из-за их гибкости ( Идентификатор PDB: 1MDM). Красная стрелка указывает на особенно длинный неупорядоченный сегмент, который адаптируется к структуре ДНК ( f ). Слева: кристаллическая структура ядерной частицы нуклеосомы Xenopus laevis (PDB id: 1AOI).Обертывающая ДНК включает две копии ядерных гистонов h3A, h3B, h4 и h5. Справа: отдельные виды восьми гистонов в геометрии внутри нуклеосомной частицы с одинаковыми цветами и неупорядоченными сегментами, показанными в виде непрозрачных лент.
В комплексе 1: 1 Dsup прочно связан с ДНК через различные структурные области (рис. 8a). Плотность силовых линий, в которых Dsup располагается вокруг ДНК, предполагает связь с очень сильными электрическими полями (рис. 8b). В комплексе 2: 1 присутствие второй молекулы Dsup вводит пространственное ограничение в динамическую эволюцию обоих белков, которое отмечается в общем меньшем интерфейсе взаимодействия по сравнению с тем, что обнаруживается в комплексе 1: 1 (сравните рис.8а, в). Несмотря на это, протяженные структурные области как DsupA, так и DsupB обнаруживают непосредственную близость к ДНК. Электрическое поле вокруг ДНК в этом комплексе 2: 1 (рис. 8d) демонстрирует особенности, аналогичные тем, которые обнаруживаются в комплексе 1: 1, хотя можно было ожидать, что присутствие второй молекулы Dsup будет влиять на электрическое поле. В обоих комплексах второй из строго консервативных мотивов в Dsup (сегмент 262–275) и Dsup-подобный гораздо ближе к ДНК, чем первый (сегмент 90–100), особенно в комплексе 1: 1 (рис.8a) и в DsupB в комплексе 2: 1 (рис. 8c), как уже указывалось в предыдущих разделах результатов. Большая подвижность неупорядоченной структуры могла бы в принципе допускать электростатическое притяжение между несколькими сегментами обеих молекул Dsup, что затем было бы замечено в возникновении многих силовых линий электрического поля между ними. Однако окончательная геометрия на рис. 8d показывает очень небольшое количество силовых линий электрического поля между двумя молекулами Dsup. Кажется очевидным, что однородный отрицательный электрический характер ДНК приводит все взаимодействие к электростатическому преобладанию эффектов ДНК-белок над возможными эффектами белок-белок.
Вышеупомянутая способность IDPs приспосабливать свою структуру к структуре своих партнеров очевидна в локальной геометрии, принятой сегментами Dsup ближе к ДНК в нескольких сайтах взаимодействий (Supplementary Fig. 2). Неупорядоченный белковый каркас приспосабливает свою локальную структуру к ДНК, которая, в свою очередь, может изгибаться, изгибаться или принимать другие формы. В этой связи необходимо подчеркнуть, что наши MD-траектории должным образом описывают гибкость ДНК, быстро изменяя идеальную модель B-ДНК, используемую для исходной геометрии.Не только в конечных структурах (рис. 8a, c и дополнительный рис. 2), но и во всем моделировании (дополнительные рис. 1, 3 и 4) молекула ДНК фактически принимает изогнутую и изогнутую геометрию независимо от его взаимодействие с белком. В свою очередь, чрезвычайная гибкость неупорядоченного Dsup позволяет ему регулировать флуктуирующие конформации ДНК, потому что их взаимодействие определяется электростатическими эффектами, а не конкретной геометрией. Эта структурная перестройка включает сайты взаимодействия, которые охватывают значительно большие сегменты белка.Во всех этих сайтах взаимодействие в основном осуществляется положительно заряженными боковыми цепями остатков лизина и аргинина, которые ориентированы в сторону ДНК (дополнительный рис. 2) из-за отрицательных зарядов, которые равномерно выстилают поверхность ДНК. Электростатическая комплементарность между большей частью поверхности Dsup и всей поверхностью ДНК тем более очевидна, когда обе молекулы приближаются друг к другу (дополнительные рисунки 3 и 4). Изменения, происходящие при уменьшении минимального расстояния между Dsup и ДНК (рис.6) намекают на важную роль электростатики в объединении больших сегментов двух молекул (дополнительные рисунки 3 и 4). Эти изменения предполагают, что возможная защитная роль Dsup в защите ДНК от внешних воздействий должна быть следствием электростатического притяжения, которое вызывает слияние поверхностей обеих молекул, что также отражается в генерации интенсивных электрических полей (рис. 3 и 8). Гибкость Dsup позволяет этому экранированию действовать независимо от конформации ДНК, так что со структурной точки зрения его предполагаемая защитная роль может быть одинаково сыграна либо на свободной ДНК, либо на кольцевой ДНК в нуклеосомах.
Наконец, наличие вторичной структуры во всех моделях MD в основном ограничивается переходными небольшими элементами (дополнительная таблица S1). Эти элементы представляют собой, с одной стороны, несколько небольших (не более 4 остатков) α- и 3 10 -спиралей в основном на участках последовательности ~ 100–125, ~ 160–170, ~ 270–290 и ~ 400–425 , а с другой стороны, несколько коротких (3 остатка) β-цепей, разбросанных по всей последовательности. Что касается результатов МД, область ~ 140–220, для которой мы обнаружили, что модели, основанные на локальной гомологии, были предсказаны (рис.1c) и что предыдущий анализ in silico предсказал структурированную альфа-спиральную область 14, 19 , не показывает тенденций к более высокому содержанию вторичной структуры, чем остальные области.
Белок-супрессор повреждений тихоходок связывается с нуклеосомами и защищает ДНК от гидроксильных радикалов.
AbstractТихоходки, также известные как водяные медведи, — животные, способные выжить в экстремальных условиях. Тихоходка Ramazzottius varieornatus содержит уникальный ядерный белок, называемый Dsup, для подавления повреждений, который может повышать устойчивость человеческих клеток к повреждению ДНК в таких условиях, как ионизирующее излучение или обработка перекисью водорода, которые генерируют гидроксильные радикалы.Здесь мы обнаруживаем, что R. varieornatus Dsup является нуклеосомо-связывающим белком, который защищает хроматин от гидроксильных радикалов. Более того, ортолог Dsup из тихоходки Hypsibius instanceplaris аналогичным образом связывается с нуклеосомами и защищает ДНК от гидроксильных радикалов. Поразительно, что консервативная область в белках Dsup демонстрирует сходство последовательностей с нуклеосомо-связывающим доменом белков HMGN позвоночных и функционально важна для связывания нуклеосом и защиты гидроксильных радикалов. Эти данные предполагают, что Dsup способствует выживанию тихоходок в различных условиях с помощью прямого механизма, который включает связывание с нуклеосомами и защиту хромосомной ДНК от гидроксильных радикалов.
Многие научные публикации, созданные UC, находятся в свободном доступе на этом сайте из-за политики открытого доступа UC. Сообщите нам, насколько этот доступ важен для вас.
Основное содержаниеЗагрузить PDF для просмотраПросмотреть больше
Больше информации Меньше информации
Закрывать
Введите пароль, чтобы открыть этот PDF-файл:
Отмена ОК
Подготовка документа к печати…
Отмена
«Подавитель повреждений» протеин защищает очаровательных тихоходок… и человеческие клетки, тоже
Ученые недавно расшифровали ключевой ингредиент в арсенале сверхспособностей тихоходок, выяснив, как уникальный белок в любимых всеми микроскопических водных медведях действует как барьер против вредного излучения.
Хотя и крошечные, тихоходок известны своей выносливостью. Они могут выдерживать экстремальные условия, которые убивают большинство форм жизни, включая воздействие холода, палящего зноя, вакуума и смертельной радиации космоса .
Но какие химические секреты делают тихоходки почти неуязвимыми? Чтобы ответить на этот вопрос, исследователи внимательно изучили соединение, обнаруженное только у тихоходок: так называемый белок-супрессор повреждений, или Dsup.
Ранее было обнаружено, что защитные свойства этого белка распространяются не только на тихоходок; при добавлении к человеческим клеткам Dsup защищает от повреждения рентгеновскими лучами. И теперь ученые обнаружили, как Dsup связывается со структурами хромосом и защищает ДНК от вредного воздействия радиации, сообщили исследователи в новом исследовании.
Связано: 8 причин, почему мы любим тихоходок
«Мы думали, что этот удивительный белок в экстремальном организме может рассказать нам что-то новое, чего мы не получим из обычных белков», — сказал соавтор исследования Джеймс. Кадонага, профессор Отделения биологических наук Калифорнийского университета в Сан-Диего.
Хотя тихоходки могут показаться неразрушимыми , им нужна вода, чтобы быть активными и размножаться.В отсутствие воды они впадают в состояние приостановки жизнедеятельности, называемое состоянием тунца, вытесняя влагу из своих тел и пребывая в высохшем подвешенном состоянии до тех пор, пока не вернутся более благоприятные условия.
Будучи тунцами, тихоходки невосприимчивы к большинству форм вреда и даже могут быть воскрешены через десятилетия, возможно, даже после того, как они проведут время на Луне. Тысячи бочек, возможно, были разбросаны по поверхности Луны после того, как израильский лунный спускаемый аппарат «Берешит» (на борту которого находились иссушенные водяные медведи) разбился 11 апреля во время неудачной попытки приземления.При определенных условиях, если они выживут при аварийной посадке, эти лиофилизированные тихоходки все еще могут вернуться к жизни, как ранее сообщал Live Science .
Кажется неразрушимым
Некоторые белки, которые позволяют тихоходкам оживать после высыхания, встречаются у других организмов, но Dsup является исключительным для водяных медведей. И хотя предыдущие исследования показали, что этот белок делает человеческие клетки устойчивыми к рентгеновскому излучению, механизмы того, как это делает Dsup, были неопределенными.
В новом исследовании исследователи обнаружили, что Dsup связывается со структурой, называемой хроматином, пакетом, который содержит длинные нити ДНК клетки в плотной упаковке, сказал Кадонага Live Science.
«Мы обнаружили, что он связывается с хроматином. Затем мы спросили:« Как он делает его устойчивым к рентгеновским лучам? »», — сказал он.
Когда клетки подвергаются воздействию рентгеновских лучей, молекулы воды расщепляются и образуют высокореактивные частицы кислорода и водорода, называемые гидроксильными радикалами ; Согласно исследованию, эти радикалы могут повредить ДНК внутри клеток.
«Мы подумали:« Почему бы нам просто не посмотреть, может ли Dsup защитить ДНК от гидроксильных радикалов? » И ответ — да, может », — объяснил Кадонага. Высокоэнергетический Dsup имеет облачную структуру; Облако окружает хроматиновую оболочку ДНК, блокируя гидроксильные радикалы и не позволяя им разрушать клеточную ДНК, сообщили исследователи.
«Теперь, когда мы знаем, как это работает, это ступенька к потенциальному использованию его в практических приложениях», — сказал Кадонага.
Собирая по кусочкам, как Dsup функционирует на все более точных уровнях, ученые могут затем использовать его в качестве схемы для создания других типов белков — «улучшенных версий Dsup», которые еще более эффективны для защиты клеток от повреждения ДНК. Кадонага сказал.Эти новые белки, вероятно, не будут использоваться для производства людей, защищенных от радиации, но они могут повысить устойчивость культивируемых клеток, которые используются для выращивания фармацевтических препаратов, добавил он.
«У вас могут быть более прочные клетки, более долгоживущие клетки. Это может быть случаем для помещения в эту клетку некоторой формы Dsup», — сказал он.
Результаты были опубликованы в Интернете во вторник (1 октября) в журнале eLife .
Первоначально опубликовано на Live Science .
Хотите больше науки? Вы можете получить 5 выпусков журнала «Как это работает» нашего партнера за 5 последних удивительных научных новостей. (Изображение предоставлено Future plc)Это медведь, это свинья, это тихоходка! Супергеройский белок Dsup защищает хроматин от вреда
Некоторые супергерои носят яркие костюмы и накидки, которые вы не могу не заметить, но есть и скромная тихоходка; микроскопический чудо-зверь с поразительной способностью выживать в экстремальных условиях температуры, радиация и космический вакуум! С несколькими секретными идентификаторами, включая «Водяной медведь» и «моховой поросенок», дискретное беспозвоночное сохранило ключ к его супер-навыки выживания были скрыты до недавнего времени.
Белок-супрессор повреждений (Dsup) одного вида тихоходка может защитить человеческие клетки от повреждающих рентгеновских лучей, и лаборатория Джеймса Кадонага (Калифорнийский университет, Сан-Диего) была исследуем, как это работает. Талантливая команда сгенерировал версии Dsup с тегами FLAG и His6 и изучил, как он защищает невинный хроматин от злодеев свободных радикалов.
Они обнаружили, что Dsup:
- Предпочтительно связывается с отдельными нуклеосомами
над изолированной ДНК, независимо от последовательности, в анализах сдвига подвижности геля
- Две молекулы Dsup, вероятно, связываются с каждой нуклеосомы, потому что на геле появляются две отдельные полосы
- Не влияет на способность плазмиды
ДНК для образования сверхспиральных структур при индуцированной сборке нуклеосом
- При разделении вновь образованных нуклеосом с MNase при переваривании и прогоне на геле, Dsup был обнаружен в основном в моно- и динуклеосомы
- Не мешает естественным нуклеосомам связывающий белок, гистон h2 в образовании нуклеосом
- очень похож на белок в другом тихоходки, и оба белка связываются с мононуклеосомами и защищают хроматин от деградации гидроксильными радикалами
Они также обнаружили, что Dsup содержит ДНК-связывающую последовательность это очень похоже на белок, связывающий нуклеосомы, обнаруженный у людей. (HMGN), и он необходим для связывания хроматина.
Может показаться, что мы раскрыли все тайны Dsup, но тихоходки хранят по крайней мере еще один секрет о своей сверхспособности: HMGN встречается только у позвоночных, так как же его гомолог оказался в воде? геном медведя? Пока мы ждем продолжения, биоинженеры могут начните использовать Dsup для защиты ценных линий клеток человека, и вы сможете выкопать подробности этой саги в eLife , октябрь 2019.
Секрет выносливости тихоходок раскрыт с помощью суперкомпьютерного моделирования
Роуэн Хупер
Тихоходки необычайно выносливы
СТИВ ГШМЕЙССНЕР / НАУЧНАЯ ФОТОБИБЛИОТЕКА
Самое стойкое животное из известных науке — тихоходка — раскрывает свои секреты: первая работа на атомном уровне посвящена изучению того, как животное переживает экстремальный стресс.
Тихоходки — микроскопические восьминогие животные, которых иногда называют водяными медведями. Они обитают во мхе по всему миру, что также послужило поводом для причудливого и откровенно млекопитающецентрического названия «Моховой поросенок».
Реклама
Ни одно млекопитающее не выдержит того, что терпит тихоходка. Под воздействием экологического стресса, такого как обезвоживание или экстремальные температуры, они сжимаются до состояния «бочки», в котором их метаболизм практически останавливается.В этом состоянии они могут выжить без воды в течение десятилетий, выдерживают высокие дозы гамма- и рентгеновского излучения и выдерживают температуры от -272 ° C до 150 ° C. Они также пролетели 10 дней в космическом вакууме.
У большинства других организмов стрессы такого рода разрушают ДНК в клетках, но тихоходки имеют белок-супрессор повреждений (Dsup), который каким-то образом защищает ДНК. Теперь Марина Мингес-Тораль и ее коллеги из Центра биотехнологии и геномики растений в Мадриде, Испания, провели симуляцию взаимодействия между Dsup и ДНК, которая предлагает объяснение.
Команда смоделировала систему из двух молекул Dsup и ДНК, состоящую из более чем 750 000 атомов, что потребовало «дней и дней» на суперкомпьютере. «Уравнения движения должны быть решены для каждого из этих атомов 50 миллионов раз, чтобы получить симуляцию продолжительностью 100 наносекунд», — говорит Мингес-Тораль.
Моделирование исследователями всех атомов в белке и всех их электростатических взаимодействий показывает, что белок «внутренне неупорядочен» и очень гибок, и, кажется, способен регулировать свою структуру, чтобы точно соответствовать форме ДНК.
«Наше исследование показывает, что электрические эффекты, лежащие в основе притяжения положительно-отрицательного заряда, определяют динамику структурных изменений Dsup при его взаимодействии с ДНК», — говорит Мингес-Тораль. «Мы считаем, что это электрическое экранирование имеет первостепенное значение для защиты ДНК от радиации».
Точное выяснение того, как тихоходки переносят такие крайности, может быть полезным по нескольким причинам. «Сейчас мы активно работаем над стабилизацией фармацевтических препаратов и созданием устойчивых к стрессу сельскохозяйственных культур», — говорит Томас Бутби, работающий с тихоходками из Университета Вайоминга.
Другие возможности включают лечение рака, а также футуристические приложения, такие как спячка человека и космические путешествия — например, люди, отправляющиеся на Марс, могут быть изменены, чтобы стать более устойчивыми к радиации. В лаборатории клетки почек человека были генетически модифицированы для экспрессии Dsup тихоходок, и эти клетки показали снижение на 40-50% повреждений ДНК, вызванных рентгеновскими лучами.
Следующим этапом будет изменение всех клеток в другом организме, и лабораторные фавориты, такие как аскариды и плодовая муха, выглядят как хорошие кандидаты, говорит Бутби.Однако Мингес-Торал и его коллега Луис Пасиос подчеркивают, что мы не знаем, почему Dsup эволюционировал, и нам нужно выяснить это, прежде чем мы начнем думать о модификации целых организмов.